

Система учета научной деятельности (ASSA) |


|
Лаборатория механизмов клеточной дифференцировки (т.26)Отдел биологии клетки
Научные результаты Сотрудники О Подразделении 1. Основное направление фундаментальных и фундаментально-ориентированных исследований
а)исследование оогенеза б)исследование формирования глаза дрозофилы в)исследование процесса формирования дорзальных выростов хориона (ДВХ). ДВХ представляют собой трубки, обеспечивающие дыхание развивающегося эмбриона. ДВХ являются перспективной моделью для исследования тубуллогенеза - процесса лежащего в основе формирования многих типов органов эукариот
2. Задачи, решаемые в настоящее время в рамках базового бюджетного проекта Бюджетный проект VI.47.1. “Механизмы контроля молекулярно-генетических систем и процессов. Нанобиоинженерия” (координатор программы. д.б.н., проф. Меркулова Т.И.). Проект направлен на решение одной из ключевых проблем молекулярной генетики – изучение регуляторного кода ДНК эукариот и выяснение механизмов определяющих характер транскриптома эукариотической клетки. Лаборатория механизмов клеточной дифференцировки отвечает за блок «Идентификация генов-мишеней транскрипционных факторов, участвующих в контроле развития органов дрозофилы». Задачи этого блока: анализ транскриптома ряда органов D. melanogaster и выявление генов-мишеней транскрипционного фактора GAGA с использованием гипоморфных мутаций по гену, кодирующему этот белок в геноме с помощью комплекса экспериментальных и компьютерных методов. Бюджетный проект VI.60.1.3. «Генетические и эпигенетические механизмы, регулирующие дифференцировку, трансдифференцировку и перепрограммирование» (координатор – д.б.н. О.Л.Серов). Лаборатория механизмов клеточной дифференцировки отвечает за блок «Клеточные и молекулярные механизмы гонадогенеза у дрозофилы» Задачи этого блока: изучение миграции половых клеток в эмбриогенезе дрозофилы и анализ особенностей дифференцировки половых клеток в зрелых гонадах, выявления механизмов их регуляции, а также факторов, нарушающих их протекание.
3. Прикладные разработки Прикладных разработок нет.
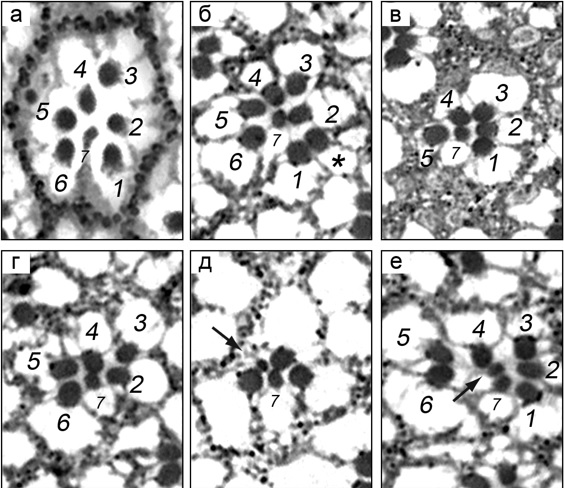
4. Иллюстрированное описание лучших результатов, полученных подразделением за последние 5 лет Установлено, что продукт гена Trihorax-like (Trl) – транскрипционный фактор GAGA является важным регулятором развития глаза дрозофилы. Ранее было известно, что нарушение экспрессии гена Trl приводит к нарушениям структуры глаза. Детально проанализировав характер нарушений, мы установили, что ген Trl является важным регулятором процессов дифференцировки нескольких типов клеток формирующегося глаза. Так, было выявлена роль ТФ GAGA в контроле формирования фотонейронов (рис. 1), конусных и пигментных клеток (рис. 2) омматидия.
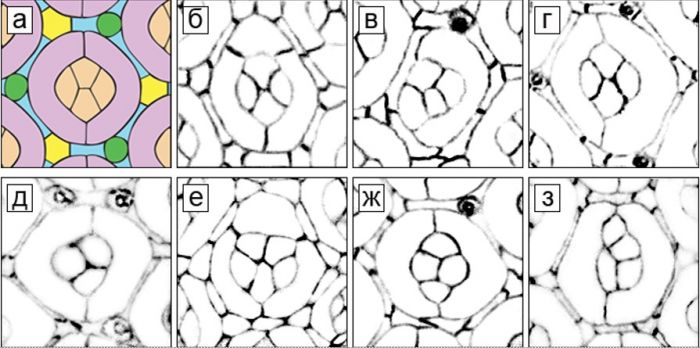
Рисунок 2. Нарушения количества и расположения конусных и первичных пигментных клеток у гетерозигот Trl en82/Trl R85 и Trl 362/Trl R85. (а) Схема расположения клеток на апикальной поверхности глазо-антеннального имагинального диска 42-х часовых куколок дрозофилы дикого типа. Коричневым цветом отмечены 4 конусные клетки (КК), сиреневым ‑ 2 первичные пигментные клетки (ППК), голубым – вторичные пигментные клетки, желтым – третичные пигментные клетки, зеленым – щетинки. (б‑з) Омматидии Trl-мутантов. В них присутствуют: (б) дополнительная ППК; (в) всего одна ППК; (г) 2 ППК неодинакового размера со смещенными контактами; (д) 3 КК; (e) 5 КК; (ж) 4 КК с измененной ориентацией контактов; (з) 4 КК с нарушением контактов между ними. Окрашивание антителами к белку Armadillo (Arm).
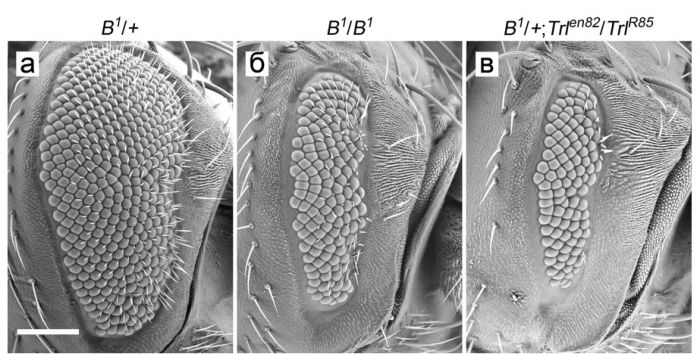
В ходе выяснения механизмов действия GAGA было установлено, что он участвует в регуляции двух ключевые регуляторов развития глаза генов lozenge (lz) и Bar. В них обнаружены сайты связывании ТФ GAGA. Кроме того продемонстрировано генетическое взаимодействие между этими генами и геном Trl (рис. 3). Установлено, что экспрессия гена lz нарушается на фоне снижения количества ТФ GAGA.
Рисунок 3. Усиление проявления мутантного фенотипа дупликации B1 гена BarH1 на фоне мутаций по гену Trl. (а) Глаз самки B1/+. (б) Глаз самки B1/B1. (в) Глаз самки B1/+; Trl en82/Trl R85. Масштаб: 100 мкм.
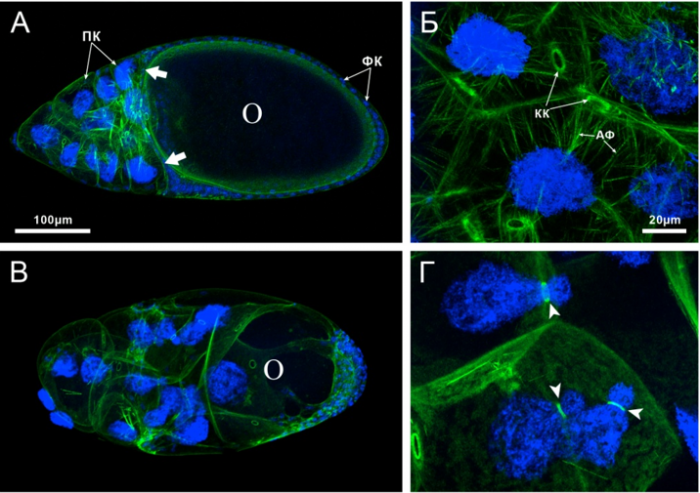
Ранее нами было установлено, что недостаток ТФ GAGA приводит к множественным нарушениям процесса оогенеза у дрозофилы. У мутантов по гену наблюдаются множественные нарушения в функционировании разных типов клеток. Так уменьшение экспрессии гена приводит к нарушению актинового цитоскелета в питающих клетках. В них практически полностью отсутствуют цитоплазматические актиновые филаменты, удерживающие большие полиплоидные ядра клеток вдали от кольцевых канальцев на стадии быстрого транспорта (рис. 4). В результате кольцевые канальцы блокируются этими ядрами и ток цитоплазмы прекращается. Это приводит к тому, что недополучившие необходимые для развития вещества ооцита прекращают развитие и гибнут.
Рисунок 4. Нарушения, наблюдаемые в яйцевых камерах сильных Trl мутантов. Яйцевые камеры 11 стадии развития, окрашенные фаллоидином (зеленый) и DAPI (синий). (А,Б) Яйцевые камеры самок дикого типа и (В,Г) Trl мутантов. У Trl мутантов цитоплазматические актиновые филаменты отсутствуют, ядра питающих клеток блокируют кольцевые каналы. Нарушена миграция фолликулярных клеток, большинство фолликулярных клеток у мутантов сосредоточено в заднем конце ооцита. ПК – питающие клетки, ФК – фолликулярные клетки, КК – кольцевые каналы, АФ – цитоплазматические актиновые филаменты, О – ооцит. Стрелками показаны центрипитальные клетки.
Кроме того у мутантов наблюдается нарушения в миграции ряда соматических клеток в ходе развития яйцевых камер. Для выяснения причин возникновения наблюдаемых дефектов нами был сделан полногеномный микрочиповый анализ экспрессии в яйцевых камерах в норме и мутантов (рис. 5). Был установлен набор генов изменяющих экспрессию на фоне снижения белка GAGA. Среди них были отобраны гены, кодирующие актин-связывающие белки и меняющие экспрессию у мутантов. Был проведен анализ экспрессии этих генов у мутантов с помощью количественного ПЦР, а также проверено наличие генетического взаимодействия между геном Trl и предполагаемыми генами-мишенями. В результате было установлено, ТФ GAGA необходим для экспрессии таких генов актин-связывающих белков как: chic, MSP-300, tsr, jar и нескольких других. Вероятно, нарушения в количестве этих актин-связывающих белков и приводит к дефектам в формировании актинового цитоскелета у мутантов и нарушению миграции соматических клеток.
5. Задачи, планируемые на перспективу
Березина Дарья Андреевна [лаборант] Брусенцов Илья Иванович [научный сотрудник] Ваулин Олег Викторович [научный сотрудник] Голыгина Вероника Вилорьевна [научный сотрудник] Карагодин Дмитрий Александрович [младший научный сотрудник] Романюк Жанна Владимировна [старший лаборант] Чадов Борис Федорович [научный консультант] Шарахова Мария Владимировна [научный сотрудник] Шилова Валентина Михайловна [старший лаборант] Бывшие сотрудникиАрискина Галина ВасильевнаБаттулина Надежда Викторовна Брошков Андрей Дмитриевич Зиневич Елена Викторовна Лактионов Петр Павлович Огиенко Анна Александровна Омелина Евгения Сергеевна Пиндюрин Алексей Валерьевич Похолкова Галина Витальевна Соловьев Виктор Владимирович Федорова Елена Владимировна Федорова Нина Борисовна Чадова Евгения Васильевна СовместителиБухарина Анастасия Владимировна [лаборант]Выберите слайдером нужный промежуток, и список ниже будет содержать записи только нужного периода: 01 02 03 04 05 06 07 08 09 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 Публикации Монографии Конференции Гранты Научное руководство Патенты
|


| 2025 | Chromosomal polymorphism in natural populations of Chironomus borokensis Kerkis, Filippova, Shobanov, Gunderina et Kiknadze, 1988 (Diptera, Chironomidae) Golygina VV, Broshkov AD, Ermolaeva OV COMP CYTOGENET, 2025, 19:51-74 |
| Chironomus sp. J Kiknadze, 1991 - the elusive species of Chironomus plumosus (Linnaeus 1758) sibling-species group (Diptera, Chironomidae). Golygina V.V. COMP CYTOGENET, 2025, 19: 209-219 |
|
| 2024 | The chromosome-scale genome assembly for the West Nile vector Culex quinquefasciatus uncovers patterns of genome evolution in mosquitoes Ryazansky S. S., Chen C., Potters M., Naumenko A. N., Lukyanchikova V., Masri R. A., Brusentsov, II, Karagodin D. A., Yurchenko A. A., Dos Anjos V. L., Haba Y., Rose N. H., Hoffman J., Guo R., Menna T., Kelley M., Ferrill E., Schultz K. E., Qi Y., Sharma A., Deschamps S., Llaca V., Mao C., Murphy T. D., Baricheva E. M., Emrich S., Fritz M. L., Benoit J. B., Sharakhov I. V., McBride C. S., Tu Z., Sharakhova M. V. BMC BIOL, 2024, V. 22, № 1. P. 16 |
| Ontogenes in Drosophila Melanogaster and a Model of Speciation Chadov B.F., Fedorova N.B. J.Evolutionary Science, 2024, 1(3):35-58 |
|
| Genetic diversity of populations of Simulium reptans s.l. (Diptera: Simuliidae) in
the territory of Russia and Northern Kazakhstan Vaulin O.V., Petrozhitskaya L.V., Novgorodova T.A. ZOOTAXA, 2024, 5543 (3): 329–342 |
|
| Cytogeography of the Polyploid Complex Bassia prostrata s. l. (Chenopodiaceae) Based on Genome Size Analysis and PCR-RFLP cpDNA T. V. Pankova, M. N. Lomonosova, O. V. Vaulin, A. Yu. Korolyuk, E. A. Korolyuk, D. N. Shaulo, B. Osmonali RUSS J GENET+, 2024, V.60, No 3, pp. 274–287 |
|
| 2023 | The complex role of transcription factor GAGA in germline death during Drosophila spermatogenesis: transcriptomic and bioinformatic analyses Fedorova S, Dorogova NV, Karagodin DA, Oshchepkov DY, Brusentsov II, Klimova NV, Baricheva EM. PeerJ, 2023, PeerJ. 2023 Jan 9;11:e14063 |
| The Multiple Mitotic Roles of the ASPM Orthologous Proteins: Insight into the Etiology of ASPM-Dependent Microcephaly Alyona V. Razuvaeva, Lucia Graziadio, Valeria Palumbo, Gera A. Pavlova, Julia V. Popova, Alexey V. Pindyurin, Silvia Bonaccorsi, Maria Patrizia Somma, Maurizio Gatti. Cells, 2023 |
|
| Formica gagatoides Ruzsky, 1904, and Siberian F. kozlovi Dlussky, 1965 (Hymenoptera: Formicidae); Two or One Species? Chesnokova, S.V.; Vaulin, O.V.; Zhigulskaya, Z.A.; Novgorodova, T.A. Diversity, 2023, Diversity 2023, 15, 686. |
|
| The misregulation of mitochondria-associated genes caused by GAGA-factor lack promotes autophagic germ cell death in Drosophila testes Dorogova NV, Fedorova SA, Bolobolova EU, Baricheva EM. GENETICA, 2023 |
|
| Ontogenes and Their Role in Cellular
Construction Chadov, B. F., and Fedorova, N.B. Advances in Bioscience and Biotechnology, 2023, v 14, p 49-73 |
|
| Mutation in Ontogene and Emergence of
Secondary Chromosome Damages in Drosophila Germline Cells Chadov, B.F., and Fedorova, N.B. Advances in Bioscience and Biotechnology, 2023, v14, p 379-398 |
|
| Genetic Analysis of the Characters of Intraspecific Similarity. Chadov, B. F., and Fedorova, N. B. Med Res Archives, 2023, Vol. 8, Chapter 3: 20–32 |
|
| New Class of Genes in D. melanogaster (Conditional Mutations, Ontogenes, and Biological Role of Ontogenes). Chadov, B.F., and Fedorova, N.B. Japan Journal of Research, 2023, 4(6): 1-8 |
|
| Patterns of genetic differentiation imply distinct phylogeographic history of the mosquito species Anopheles messeae and Anopheles daciae in Eurasia Brusentsov I. I., Gordeev M. I., Yurchenko A. A., Karagodin D. A., Moskaev A. V., Hodge J. M., Burlak V. A., Artemov G. N., Sibataev A. K., Becker N., Sharakhov I. V., Baricheva E. M., Sharakhova M. V. MOL ECOL, 2023, P: 1–17 |
|
| Phylogenomics revealed migration routes and adaptive radiation timing of Holarctic malaria mosquito species of the Maculipennis Group Yurchenko A. A., Naumenko A. N., Artemov G. N., Karagodin D. A., Hodge J. M., Velichevskaya A. I., Kokhanenko A. A., Bondarenko S. M., Abai M. R., Kamali M., Gordeev M. I., Moskaev A. V., Caputo B., Aghayan S. A., Baricheva E. M., Stegniy V. N., Sharakhova M. V., Sharakhov I. V. BMC BIOL, 2023, V. 21, № 1. P. 63 |
|
| 2022 | Genetic control of kinetochore-driven microtubule growth in Drosophila mitosis Popova JV, Pavlova GA, Razuvaeva AV, Yarinich LA, Andreyeva EN, Anders AF, Galimova YA, Renda F, Somma MP, Pindyurin AV, Gatti M. Cells, 2022, 11(14), 2127 |
| Ontogenes and Chromosome Nondisjunction in the D. melanogaster Meiosis Boris F. Chadov, Nina B. Fedorova Advances in Bioscience and Biotechnology, 2022, Vol.13, № 8, p. 317-335 |
|
| Chromosomal polymorphism in natural populations of Chironomus sp. prope agilis Kiknadze, Siirin, Filippova et al., 1991 (Diptera, Chironomidae) Golygina V.V. COMP CYTOGENET, 2022, 16 (4): 243-252 |
|
| Molecular genetics of Simulium (Wilhelmia) equinum (Linnaeus) (Diptera: Simuliidae) from West Siberia. Vaulin O.V., Petrozhitskaya L.V., Novgorodova T.A. ZOOTAXA, 2022, Aug 19;5175(5):548-558. |
|
| 2021 | Ontogenes and the Genetics of Intraspecific Similarity Chadov B.F., Fedorova N.B. Med Res Archives, 2021, v. 9, n. 4. |
| Ontogenes and the Paradox of Homologous
Pairing B. F. Chadov, N. B. Fedorova Advances in Bioscience and Biotechnology, 2021, V 12, PP 1-9 |
|
| Недостаток белка GAGA у мутантов Trl вызывает массовую клеточную гибель в спермато- и оогенезе дрозофилы Дорогова Н.В., Зубкова А.Е., Федорова Е.В., Болоболова Е.У., Баричева Е.М., Vavilov journal of genetics and breeding, 2021, Т. 25, №3, С. 292 |
|
| New Cytogenetic Photomap and Molecular Diagnostics for the Cryptic Species of the Malaria Mosquitoes Anopheles messeae and Anopheles daciae from Eurasia Artemov G.N. Fedorova V.S. Karagodin D.A. Brusentsov I.I. Baricheva E.M. Sharakhov I.V. Gordeev M.I. Sharakhova M.V Insects, 2021, V. 12, #9, P. 835 |
|
| Yurchenko A.A., Karagodin D.A., Masri R.A., Smith R.C., Gordeev M.I., Sharakhova M.V. The new Internal Transcribed Spacer 2 diagnostic tool clarifies the taxonomic position and geographic distribution of the North American malaria vector Anopheles punctipennis Hodge J.M. Yurchenko A.A. Karagodin D.A. Masri R.A. Smith R.C. Gordeev M.I. Sharakhova M.V. MALARIA J, 2021, V. 20, #1, P. 141 |
|
| A Gene-Based Method for Cytogenetic Mapping of Repeat-Rich Mosquito Genomes Masri R.A. Karagodin D.A. Sharma A. Sharakhova M.V. Insects, 2021, V. 12, #2, P. 138 |
|
| Physical Mapping of the Anopheles (Nyssorhynchus) darlingi Genomic Scaffolds Rafael M.S. Bridi L.C. Sharakhov I.V. Marinotti O. Sharakhova M.V. Timoshevskiy V. Guimarães-Marques G.M. Santos V.S. da Silva C.G.N. Astolfi-Filho S. Tadei W.P. Insects, 2021, V. 12, #2, P. 164 |
|
| Маркерные последовательности ДНК для идентификации известных и выявления новых видов растений и животных Ваулин О.В. Захаров И.К. Письма в Вавиловский журнал генетики и селекции, 2021, Т. 7, №3, С. 119 |
|
| Author Correction: Improved reference genome of the arboviral vector Aedes albopictus Palatini U. Masri R.A. Cosme L.V. Koren S. Thibaud-Nissen F. Biedler J.K. Krsticevic F. Johnston J.S. Halbach R. Crawford J.E. Antoshechkin I. Failloux A.B. Pischedda E. Marconcini M. Ghurye J. Rhie A. Sharma A. Karagodin D.A. Jenrette J. Gamez S. Miesen P. Masterson P. Caccone A. Sharakhova M.V. Tu Z. Papathanos P.A. Van Rij R.P. Akbari O.S. Powell J. Phillippy A.M. Bonizzoni M. GENOME BIOL, 2021, V. 22, P. 205 |
|
| Revision of the banding sequence pool and new data on chromosomal polymorphism in natural populations of Chironomus agilis Shobanov et Djomin, 1988 (Diptera, Chironomidae) Golygina V.V., Ermolaeva O.V. COMP CYTOGENET, 2021, 15 (4): 527-541 |
|
| 2020 | Potential regulatory SNPs in the ATXN7L3B and KRT15 genes are associated with gender-specific colorectal cancer risk Elena Yu Leberfarb, Arina O Degtyareva, Ilya I Brusentsov, Vladimir N Maximov, Mikhail I Voevoda, Alexander I Autenshlus, Dmitriy V Morozov, Andrey V Sokolov, Tatiana I Merkulova PERS MED, 2020, Jan;17(1):43-54 |
| Полиморфизм rs2072580 T>A в перекрывающихся промоторных областях генов SART3 и ISCU, связанный с риском развития рака молочной железы А.О.Дегтярева, Е.Ю.Леберфарб, Е.Г.Ефимова, И.И.Брусенцов, А.В.Усова, Е.Л.Лушникова, Т.И.Меркулова Бюллетень экспериментальной биологии и медицины, 2020, Т.169, № 1, с.88-91 |
|
| Loss of Drosophila E3 Ubiquitin Ligase Hyd Promotes Extra Mitosis in Germline Cysts and Massive Cell Death During Oogenesis Dorogova Natalia V., Galimova Yuliya A., Bolobolova Elena Us., Baricheva Elina M., Fedorova Svetlana A. Front. Cell Dev. Biol., 2020, Front. Cell Dev. Biol., 09 November 2020 | https://doi.org/10.3389/fcell.2020.600868 |
|
| Analysis of Anopheles messeae s.l. intron gene polymorphism associated with imidacloprid resistance Oleg V. Vaulin, Dmitry. A. Karagodin, Tatiana A. Novgorodova, Viktor V. Glupov J VECTOR ECOL, 2020, V. 45 (2), P. 220-232. |
|
| GAGA Regulates Border Cell Migration in Drosophila Anna A. Ogienko , Lyubov A. Yarinich , Elena V. Fedorova , Natalya V. Dorogova , Sergey I. Bayborodin , Elina M. Baricheva and Alexey V. Pindyurin INT J MOL SCI, 2020, Int. J. Mol. Sci. 2020, 21, 7468 |
|
| Mucin-2 knockout is a model of intercellular junction defects, mitochondrial damage and ATP depletion in the intestinal epithelium M.A. Borisova, K.M. Achasova, K.N. Morozova, E.N. Andreyeva, E.A. Litvinova, A.A. Ogienko, M.V. Morozova, M.B. Berkaeva, E.Kiseleva & E.N. Kozhevnikova. SCI REP-UK, 2020, V.10, N. 21135 |
|
| INBREEDING DEPRESSION AND HETEROSIS AS THE PHENOMENA OF TWO-COMPONENT GENOME Boris F. Chadov*, PhD and Nina B. Fedorova†, PhD Adv Genet Res, 2020, V 19, PP 161-190 |
|
| Chromosome and Genome Divergence between the Cryptic Eurasian Malaria Vector-Species Anopheles messeae and Anopheles daciae Naumenko A. N., Karagodin D. A., Yurchenko A. A., Moskaev A. V., Martin O. I., Baricheva E. M., Sharakhov I. V., Gordeev M. I., Sharakhova M. V. Genes, 2020, V.11, №2 |
|
| Improved reference genome of the arboviral vector Aedes albopictus Palatini U., Masri R. A., Cosme L. V., Koren S., Thibaud-Nissen F., Biedler J. K., Krsticevic F., Johnston J. S., Halbach R., Crawford J. E., Antoshechkin I., Failloux A. B., Pischedda E., Marconcini M., Ghurye J., Rhie A., Sharma A., Karagodin D. A., Jenrette J., Gamez S., Miesen P., Masterson P., Caccone A., Sharakhova M. V., Tu Z., Papathanos P. A., Van Rij R. P., Akbari O. S., Powell J., Phillippy A. M., Bonizzoni M. GENOME BIOL, 2020, V. 21 |
|
| Genomic differentiation and intercontinental population structure of mosquito vectors Culex pipiens pipiens and Culex pipiens molestus Yurchenko A. A., Masri R. A., Khrabrova N. V., Sibataev A. K., Fritz M. L., Sharakhova M. V. SCI REP-UK, 2020, V. 10 |
|
| Фауна и экология кровососущих комаров (diptera: culicidae) государственного природного заповедника "нургуш" кировской области Панюкова Е.В., Целищева Л.Г., Пестов С.В., Колесникова А.А., Бакка С.В., Шарахова М.В Цитология, 2020, Т. 54, № 4. С. 322-240 |
|
| Structural Variation of the X Chromosome Heterochromatin in the Anopheles gambiae Complex Sharma A., Kinney N. A., Timoshevskiy V. A., Sharakhova M. V., Sharakhov I. V. Genes, 2020, V. 11, №3 |
|
| rs2072580T>A Polymorphism in the Overlapping Promoter Regions of the SART3 and ISCU Genes Associated with the Risk of Breast Cancer Degtyareva AO, Leberfarb EY, Efimova EG, Brusentsov II, Usova AV, Lushnikova EL, Merkulova TI B EXP BIOL MED+, 2020 |
|
| 2019 | Estimated resistance of the malaria mosquito Anopheles messeae s.l.
to the insecticide malathion Vaulin O.V., Karagodin D.A., Baricheva E.M.,Zakharov I.K. J VECTOR ECOL, 2019, V. 44 (1), P. 48-56 |
| О видах рода Cotoneaster Medic. (Rosaceae) в Туве Шауло Д.Н., Эрст А.С., Ваулин О.В., Анькова Т.В., Самбуу А.Д. Вестник Тувинского ГУ, 2019, Выпуск 2. Естественные и сельскохозяйственные науки, № 3 (49), 2019, C 5-19. |
|
| Ontogenes and the Problem of Speciation Boris Chadov E.V. Chadova N.B Fedorova J.Evolutionary Science, 2019, 1(1): p 33-47 |
|
| The Mutations Disturbing the Bilateral Symmetry in
Drosophila BF Chadov and NB Fedorova SCIOL Genetic Science, 2019, 2: p 139-152 |
|
| Personalized Approach to Assessing mRNA Expression of Histidine-Rich Glycoprotein and Immunohistochemical Markers in Diseases of the Breast Autenshlyus A. I. Golovanova A. V. Studenikina A. A. Brusentsov I. I. Proskurab A. V. Zhurakovskiya I. P. Arkhipova S. A. Sidorov S. V. Vavilin V. A. Lyakhovich V. V. Doklady Biochemistry and Biophysics, 2019, 484(5) |
|
| Epigenetic-genetic chromatin footprinting identifies novel and subject-specific genes active in prefrontal cortex neurons Fedor E. Gusev, Denis A. Reshetov, Amanda C. Mitchell, Tatiana V. Andreeva, Aslihan Dincer, Anastasia P. rigorenko, Gennady Fedonin, Tobias Halene, Maria Aliseychik, Andrey Y. Goltsov, Victor Solovyev, Leonid Brizgalov, Elena Filippova, Zhiping Weng, Schahram Akbarian, Evgeny I. Rogaev FASEB J, 2019, 33(7):8161-8173 |
|
| Promoter analysis and prediction in the human genome using sequence-based deep learning models Umarov R, Kuwahara H, Li Y, Gao X, Solovyev V BIOINFORMATICS, 2019, 35(16):2730-2737 |
|
| RNAi-mediated depletion of the NSL complex subunits leads to abnormal chromosome segregation and defective centrosome duplication in Drosophila mitosis Pavlova GA, Popova JV, Andreyeva EN, Yarinich LA, Lebedev MO, Razuvaeva AV, Dubatolova TD, Oshchepkova AL, Pellacani C, Somma MP, Pindyurin AV, Gatti M. PLOS GENET, 2019, 15(9): e1008371 |
|
| The role of Patronin in Drosophila mitosis Pavlova GA, Razuvaeva AV, Popova JV, Andreyeva EN, Yarinich LA, Lebedev MO, Pellacani C, Bonaccorsi S, Somma MP, Gatti M, Pindyurin AV BMC Mol Cell Biol, 2019, 20 (Suppl 1): 7 |
|
| 2018 | Inbreeding Depression and Heterosis: Explanation
by Two-Component Genome Composition Boris F Chadov SCIOL Genetic Science, 2018, 1:41-44 |
| Novel functional variants at the GWAS-implicated loci might confer risk to major depressive disorder, bipolar affective disorder and schizophrenia. Bryzgalov L.O., Korbolina E.E., Brusentsov I.I., Leberfarb E.Y., Bondar N.P., Merkulova T.I. BMC NEUROSCI, 2018, Apr 19;19(Suppl 1):22 |
|
| Novel Approach to Functional SNPs Discovery from Genome-Wide Data Reveals Promising Variants for Colon Cancer Risk Elena E. Korbolina, Ilja I. Brusentsov, Leonid O. Bryzgalov, Elena Yu. Leberfarb, Arina O. Degtyareva and Tatyana I. Merkulova HUM MUTAT, 2018, Jun;39(6):851-859 |
|
| The Ontogenes in Drosophila and the Problem of Fertility Chadov B.F. Annals of Infertility & Reproductive Endocrinology, 2018, V1, N1: 1004 |
|
| Conditional Mutations in Drosophila: Concept of Genes That Control Individual Development Boris F. Chadov, Eugenia V. Chadova, Nina B. Fedorova Advances in Bioscience and Biotechnology, 2018, V. 9, P. 243- 272 |
|
| ДИНАМИКА ВИДОВОГО СОСТАВА МАЛЯРИЙНЫХ КОМАРОВ
В СИБИРСКИХ ПОПУЛЯЦИЯХ, ВЫЯВЛЯЕМАЯ С ПОМОЩЬЮ
РЕСТРИКЦИОННОГО АНАЛИЗА Ваулин О.В., Карагодин Д.А., Захаров И.К., Баричева Э.М. Генетика, 2018, No 7, том 54. с. 832-842. |
|
| GAGA protein is required for multiple aspects of Drosophila oogenesis and female fertility Fedorova E.V., Dorogova N.V., Bolobolova E.U., Fedorova S.A. , Karagodin D.A., Ogienko A. A., A.S. Khruscheva A.S., Baricheva E. M. GENESIS, 2018, e23269 |
|
| New slbo-Gal4 driver lines for the analysis of border cell migration during Drosophila oogenesis Anna A. Ogienko Lyubov A. Yarinich Elena V. Fedorova Mikhail O. Lebedev Evgeniya N. Andreyeva Alexey V. Pindyurin Elina M. Baricheva CHROMOSOMA, 2018, 127:475–487 |
|
| мРНК гистидинбогатого гликопротеина в новообразованиях молочной железы Аутеншлюс АИ, Брусенцов ИИ, Маринкин ИО, Смирнова СА, Рукавишников МЮ, Ляхович ВВ Doklady Akademii Nauk, 2018, №5, т. 478, сс. 596-600 |
|
| The revision of chromosome III (EF) mapping in Chironomus plumosus (Linnaeus, 1758) group (Diptera, Chironomidae) Golygina V.V., Kiknadze I.I COMP CYTOGENET, 2018, 12(2):201-222 |
|
| DYNAMICS OF MALARIA MOSQUITO SPECIES COMPOSITION IN SIBERIAN POPULATIONS DETECTED BY RESTRICTION ANALYSIS VAULIN O.V., KARAGODIN D.A., ZAKHAROV I.K., BARICHEVA E.M. RUSS J GENET+, 2018, Том 54, № 7, Страницы 838-847 |
|
| Ultrastructural analysis of mitotic Drosophila S2 cells identifies distinctive microtubule and intracellular membrane behaviors Strunov A, Boldyreva LV, Andreyeva EN, Pavlova GA, Popova JV, Razuvaeva AV, Anders AF, Renda F, Pindyurin AV, Gatti M, Kiseleva E BMC BIOL, 2018, 16: 68 |
|
| 2017 | Protein and genetic composition of four chromatin types in Drosophila melanogaster cell lines Boldyreva L.V., Goncharov F.P., Demakova O.V., Zykova T.Y., Levitsky V.G., Kolesnikov N.N., Pindyurin A.V., Semeshin V.F., Zhimulev I.F. CURR GENOMICS, 2017, 18(2):214-226 |
| Ортогенез и дарвинизм: перспектива синтеза в свете данных по условным мутациям. Современные проблемы эволюции и экологии Чадов Б.Ф., Чадова Е.В., Федорова Н.Б Сборник Современные проблемы эволюции и экологии. Сборник материалов Всероссийской (с междунар. участием) научной конференции (Ульяновск, 30 – 31 марта 2017 г.) ХХХ Любищевские Чтения 2017, 2017, с.133-142 |
|
| Thermodynamic aspects of freeze drying: a case study of an “organic solvent-water” system. Ogienko A.G., Drebushchak V.A., Bogdanova E.G., Yunoshev A.S., Ogienko A.A., Boldyreva E.V., Manakov A.Yu. J THERM ANAL CALORIM, 2017, 127, pp 1593-1604 |
|
| Large porous particles for respiratory drug delivery. Glycine-based formulations. Ogienko A.G., Bogdanova E.G., Trofimov N.A., Myz S.A., Ogienko A.A., Kolesov B.A., Yunoshev A.S., Zubikov N.V., Manakov A.Yu., Boldyrev V.V., Boldyreva E.V. EUR J PHARM SCI, 2017, DOI: 10.1016/j.ejps.2017.05.007 |
|
| Роль гена pAbp, кодирующего цитоплазматический поли(А)- связывающий белок, в сперматогенезе Drosophila melanogaster. Болоболова Е.У., Баричева Э.М., Дорогова Н.В. Vavilov journal of genetics and breeding, 2017, 21(6):710-716. |
|
| Связь между фенотипической робастностью и средней продолжительностью жизни у Drosophila melanogaster Л.П. Захаренко, Д.В. Петровский, И.Г. Дранов, С.А. Федорова, Н.С. Юдин, А.В. Пиндюрин, Ю.М. Мошкин Vavilov journal of genetics and breeding, 2017, 2017;21(7):816-824. DOI 10.18699/VJ17.301 |
|
| МОЛЕКУЛЯРНАЯ ХАРАКТЕРИСТИКА СТАБИЛЬНЫХ И НЕСТАБИЛЬНЫХ АЛЛЕЛЕЙ ГЕНА white В ВЫСОКОМУТАБИЛЬНЫХ ЛИНИЯХ ИЗ ПРИРОДНЫХ ПОПУЛЯЦИЙ Drosophila melanogaster М.А. Волошина, Ю.А. Коромыслов, О.В. Ваулин, И.К. Захаров Генетика, 2017, № 12, Т. 53, С. 1380-1392 |
|
| A novel type of gene interaction in D. melanogaster Chadov B.F., Chadova E.V., Fedorova N.B. MUTAT RES-FUND MOL M, 2017, Biomedical Advances/Editors’ Picks, July 2017/ http://biomedical-advances.org/ep-20175-29/ PMID: 28103492 |
|
| The Genetics of Conditional Mutations and Individual Developmental Programs in D.melanogaster Chadov BF, Chadova EV, Fedorova NB SCIOL Genetic Science, 2017, V1, P. 3-21 |
|
| 2016 | Hemozoin is a product of heme detoxification in the gut of the most medically important species of the family Opisthorchiidae. Lvova M, Zhukova M, Kiseleva E, Mayboroda O, Hensbergen P, Kizilova E, Ogienko A, Besprozvannykh V, Sripa B, Katokhin A, Mordvinov V. INT J PARASITOL, 2016, Mar;46(3):147-56. doi: 10.1016/j.ijpara.2015.12.003. |
| Гены, кодирующие рибосомный белок RPL11 и 5,8S рибосомную РНК, у видов рода Chironomus: нуклеотидная изменчивость, дивергенция и хромосомная локализация Л.И. Гундерина, А.Д. Брошков, О.В. Ермолаева, В.В Голыгина. Цитология, 2016, Т. 58. № 5: 399-403 |
|
| Роль фактора GAGA в миграции примордиальных зародышевых клеток и формировании гонад дрозофила Дорогова Н.В., Хрущева А.С., Федорова Е.В., Огиенко А.А., Баричева Э.М. Онтогенез, 2016, том 47, № 1, с. 40–48 |
|
| DNA barcoding supports reclassification of Japanese Chironomus species (Diptera: Chironomidae) Natsuko I. Kondo, Ryuhei Ueno, Kako Ohbayashi, Veronika V. Golygina and Kenzi Takamura ENTOMOL SCI, 2016, doi: 10.1111/ens.12212 |
|
| Factors governing the pattern of spindle microtubule regrowth after tubulin depolymerization Pavlova G.A., Galimova J.A., Popova J.V., Munzarova A.F., Razuvaeva A.V., Alekseeva A.L., Berkaeva M.B., Pindyurin A.V., Somma M.P., Gatti M., Renda F. Цитология, 2016, 58(4), 299-303 |
|
| A simple and effective method for ultrastructural analysis of mitosis in Drosophila S2 cells Strunov A., Boldyreva L.V., Pavlova G.A., Pindyurin A.V., Gatti M., Kiseleva E. MethodsX, 2016, 3, 551-559 |
|
| Филогенетические связи между палеоарктическими видами Anopheles комплекса maculipennis (Diptera: Culicidae), установленные при использовании разных методов. Проблема консенсуса Ваулин О.В., Новиков Ю.М. Vavilov journal of genetics and breeding, 2016, № 5. Т. 20. С. 695-703. |
|
| Анализ геномного распределения сайтов связывания транскрипционных факторов GAGA и CNC в развитии Drosophila melanogaster Брусенцов И.И., Карагодин Д.А., Баричева Э.М., Меркулова Т.И. Vavilov journal of genetics and breeding, 2016, 20(6):762-769 |
|
| Выявление новых регуляторных SNP-маркеров, ассоциированных с предрасположенностью к развитию колоректального рака Леберфарб Е.Ю., Брызгалов Л.О., Брусенцов И.И., Меркулова Т.И. Vavilov journal of genetics and breeding, 2016, 20(6) |
|
| The reasons of Trithorax-like expression disturbance in Trl 3609 allele of Drosophila melanogaster Karagodin D. A., Battulina N. V., Merkulova T. I., Baricheva E. M. Doklady Biochemistry and Biophysics, 2016, Vol. 471, № 1, P. 443-446 |
|
| Genes and Ontogenes in Drosophila: The Role of RNA Forms Fedorova NB, Chadova EV, Chadov BF (2016) Genes and Ontogenes in Drosophila: The Role of RNA Forms // Transcriptomics 4: 137. DOI:10.4172/2329-8936.1000137. Transcriptomics, 2016, 4: 137. DOI:10.4172/2329-8936.1000137. |
|
| Transcriptomics: Genes and ontogenes in Drosophila Nina B Fedorova, Eugenia V Chadova and Boris F Chadov Transcriptomics, 2016, Volume 4, Issue 2 (Suppl) Page 27 |
|
| Послание XXI веку (к 35-летию со дня кончины Н. В. Тимофеева – Ресовского) Чадов Б.Ф. Эко-потенциал, 2016, №2 (14), 129-132 |
|
| Inducible DamID systems for genomic mapping of chromatin proteins in Drosophila Alexey V. Pindyurin, Ludo Pagie, Elena N. Kozhevnikova, Joris van Arensbergen, Bas van Steensel NUCLEIC ACIDS RES, 2016 |
|
| Role of GAGA factor in drosophila primordial germ cell migration and gonad development Dorogova N.V., Khrushcheva A.S, Fedorova E.V., Ogienko A.A., Baricheva E.M. RUSS J DEV BIOL+, 2016, Volume 47, Issue 1, pp 33–40 |
|
| Factors governing the pattern of spindle microtubule regrowth after tubulin depolymerization Pavlova GA, Galimova YuA, Popova YuV, Munzarova AF, Razuvaeva AV, Alekseeva AL, Berkaeva MB, Pindyurin AV, Somma MP, Gatti M, Renda F Цитология, 2016, 58: 299-303 |
|
| 2015 | Conditional mutations in Drosophila Chadov B.F., Fedorova N.B.,Chadova E.V. Atlas of Science, 2015, http://atlasofscience.org/conditional-mutations-in-drosophila |
| Ultrafine Betulin Formulation with Biocompatible Carriers Exhibiting Improved Dissolution Rate Svetlana A. Myza,b, Tatyana P. Shakhtshneidera,b, Mikhail A. Mikhailenkob, Andrey G. Ogienkoc,d, Ekaterina G. Bogdanovaa,c, Anna A. Ogienko, Svetlana A. Kuznetsovaf,g , Elena V. Boldyrevaa,b and Vladimir V. Boldyreva,b NAT PROD COMMUN, 2015, 2015 Vol. 10 No. 8 1345 - 1347 |
|
| К вопросу о создании института фьючеризма в науке Чадов Б.Ф. Эко-потенциал, 2015, № 1 (9). – С. 138–140 |
|
| О причине системности Вселенной и её частей Чадов Б.Ф. Эко-потенциал, 2015, №2 (10). С. 124-146. |
|
| ЭКСПЕРИМЕНТАЛЬНАЯ ОЦЕНКА ПРОТИВОВОСПАЛИТЕЛЬНОГО ДЕЙСТВИЯ КАЛЬЦИТА, ОБРАБОТАННОГО ПО ТИБЕТСКОЙ ТЕХНОЛОГИИ А.А. Торопова, Е.А. Лосев, А.А. Огиенко, Т.Н. Дребущак, В.В. Болдырев, Т.А. Асеева Традиционная медицина, 2015, Том 42, № 3. - С. 57 - 59 |
|
| A standard photomap of ovarian nurse cell chromosomes in the European malaria vector Anopheles atroparvus. Artemov GN, Sharakhova MV, Naumenko AN, Karagodin DA, Baricheva EM, Stegniy VN, Sharakhov IV. MED VET ENTOMOL, 2015, Vol 29, #3, P:230-237, doi: 10.1111/mve.12113. |
|
| Chromosomal organization of the ribosomal RNA genes in the genus Chironomus (Diptera, Chironomidae). Gunderina L.,Golygina V., Broshkov A. COMP CYTOGENET, 2015, V 9, №2, P. 201–220 |
|
| Conditional Mutations in Drosophila melanogaster: On the occasion of the 150th anniversary of G. Mendel's report in Brünn Boris F. Chadov, Nina B. Fedorova, Eugenia V. Chadova Mutation Research/Reviews in Mutation Research, 2015, Volume 765, July–September 2015, Pages 40–55 |
|
| Выявление и функциональный анализ регуляторных SNPs, связанных с развитием рака толстого кишечника Леберфарб Е.Ю., Брызгалов Л.О., Брусенцов И.И., Дегтярева А.О., Меркулова Т.И. Медицинская генетика, 2015, Том 14, №3 |
|
| Современная метафизика и циклическая протомодель Чадов Борис Федорович Эко-потенциал, 2015, 4(12). С.53-73 |
|
| Циклическая протомодель и феномен эволюции Чадов Борис Федорович Альманах "Эволюция", 2015, Отв. ред. Л.Е. Гринин, А.В. Коротаев. - Волгоград: Учитель, С.33-5720 |
|
| Tethering of SUUR and HP1 proteins results in delayed replication of euchromatic regions in Drosophila melanogaster polytene chromosomes Pokholkova G.V., Koryakov D.E., Pindyurin A.V., Kozhevnikova E.N., Belyakin S.N., Andreyenkov O.V., Belyaeva E.S., Zhimulev I.F. CHROMOSOMA, 2015, 124(2), 209-220 |
|
| Высокоэффективные лекарственные формы нового поколения:фармацевтические композиции сальбутамола и будесонида для использования в порошковых ингаляторах Огиенко А.Г., Богданова Е.Г., Болдырева Е.В., Трофимов Н.А., Мызь С.А., Огиенко А.А., Максимова В.Ц., Колесов Б.А., Дребущак В.А., Красников А.А., Манаков А.Ю., Болдырев В.В. Разработка и регистрация лекарственных средств, 2015, №1(10) февраль 2015 |
|
| 2014 | Критика концепции малых мутаций Иванов Юрий Николаевич Вестник семинара АНИ, 2014, №2, с. 64-70 |
| Общая теория видовых миграций Иванов Юрий Николаевич Вестник семинара АНИ, 2014, №2, с. 89-93 |
|
| Генетико-популяционные исследования плодовитости черного стрижа Apus apus L. Иванов Юрий Николаевич Вестник РУК, 2014, №5, с. 56-65 |
|
| Сравнение изменчивости видов апогамных и автогамных и видов с перекрестным оплодотворением Иванов Юрий Николаевич Вестник РУК, 2014, №5, с. 38-55 |
|
| Различие мутабильности у мелких и крупных мух в природных популяциях D.melanogaster и регуляторный смысл этого явления Иванов Юрий Николаевич Вестник РУК, 2014, №5, с. 66-77 |
|
| DNA Copy-Number Control through Inhibition of Replication Fork Progression 9. Nordman J.T., Kozhevnikova E.N., Verrijzer C.P., Pindyurin A.V., Andreyeva E.N., Shloma V.V., Zhimulev I.F., Orr-Weaver T.L. CELL REP, 2014 |
|
| Поиск новых генов-мишеней транскрипционного фактора GAGA в ходе оогенеза Drosophila melanogaster Е.С. Омелина, А.А. Коханенко Вестник ТГУ, 2014, № 1 (25). С. 132–153. |
|
| Циклическая протомодель о физических основах морали и нравственности Чадов Борис Федорович Эко-потенциал, 2014, №1 (5).С.198-220. |
|
| Формы материи в свете циклической протомодели Чадов Б.Ф. Эко-потенциал, 2014, №3 (7), 2014. С. 119-143 |
|
| СРАВНИТЕЛЬНОЕ МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКОЕ ИССЛЕДОВАНИЕ АЗИАТСКИХ ПОПУЛЯЦИЙ Drosophila mercatorum (DIPTERA; DROSOPHILIDAE) ПО ФРАГМЕНТУ ГЕНА COI И УЧАСТКУ ITS1-ITS2 ГЕНОВ pРНК Иванников А.В., Ваулин О.В., Коромыслов Ю.А., Захаров И.К. Vavilov journal of genetics and breeding, 2014, № 2, Т. 18 С. 308-314. |
|
| Expansion of Anopheles maculipennis s.s. (Diptera: Culicidae) to northeastern Europe and northwestern Asia: Causes and Consequences Yuri M Novikov, Oleg V Vaulin PARASITE VECTOR, 2014, doi:10.1186/1756-3305-7-389 |
|
| GAGA protein is essential for male germ cell development in Drosophila Дорогова Наталья Владимировна, Болоболова Елена Устиновна, Федорова Елена Владимировна, Огиенко Анна Александровна,Баричева Элина Михайловна. GENESIS, 2014, Т.52, №8, стр.738-751 |
|
| Comparative genome sequencing reveals genomic signature of extreme desiccation tolerance in the anhydrobiotic midge Oleg Gusev, Yoshitaka Suetsugu, Richard Cornette, Takeshi Kawashima, Maria D. Logacheva, Alexey S. Kondrashov, Aleksey A. Penin, Rie Hatanaka, Shingo Kikuta, Sachiko Shimura, Hiroyuki Kanamori, Yuichi Katayose, Takashi Matsumoto, Elena Shagimardanova, Dmitry Alexeev, Vadim Govorun, Jennifer Wisecaver, Alexander Mikheyev, Ryo Koyanagi, Manabu Fujie, Tomoaki Nishiyama, Shuji Shigenobu, Tomoko F. Shibata, Veronika Golygina, Mitsuyasu Hasebe, Takashi Okuda, Nori Satoh, Takahiro Kikawada NAT COMMUN, 2014, 5:4784. DOI: 10.1038/ncomms5784 |
|
| ПРИМЕНЕНИЕ СКАНИРУЮЩЕЙ ЭЛЕКТРОННОЙ МИКРОСКОПИИ В РАБОТАХ ПО ДИЗАЙНУ ЛЕКАРСТВЕННЫХ ФОРМ НОВОГО ПОКОЛЕНИЯ А. A. Огиенко, С. А. Мызь, Е. В. Болдырева, Е. Г. Зевак, А. Г. Огиенко, А. Ю. Манаков, А. И. Анчаров, Б. М. Кучумов, А. А. Красников ИЗВЕСТИЯ РАН. СЕРИЯ ФИЗИЧЕСКАЯ, 2014, том 78, № 9, с. 1107–1113 |
|
| 2013 | Долго ли осталось жить человечеству? Иванов Юрий Николаевич Вестник семинара АНИ, 2013, №1, с. 100-110 |
| Симпатрическая дивиргенция видов как воображаемое явление Иванов Юрий Николаевич Вестник РУК, 2013, №4, с 121-133 |
|
| ПРИМЕНЕНИЕ ПОРОШКОВОЙ ДИФРАКЦИИ IN SITU ДЛЯ ИССЛЕДОВАНИЯ ПРОЦЕССОВ, ПРОИСХОДЯЩИХ ПРИ ОТЖИГЕ ЗАМОРОЖЕННЫХ РАСТВОРОВ В СИСТЕМАХ С КЛАТРАТООБРАЗОВАНИЕМ В РАБОТАХ ПО СОЗДАНИЮ ВЫСОКОЭФФЕКТИВНЫХ ЛЕКАРСТВЕННЫХ ФОРМ НОВОГО ПОКОЛЕНИЯ А. Г. Огиенко, Е. В. Болдырева, А. Ю. Манаков, Е. Г. Зевак, А. А. Огиенко, С. А. Мызь, А. И. Анчаров, А. С. Юношев, М. П. Шинкоренко, А. В. Ильдяков ИЗВЕСТИЯ РАН. СЕРИЯ ФИЗИЧЕСКАЯ, 2013, том 77, № 2, с. 153–157 |
|
| Capping protein beta is required for actin cytoskeleton organisation and cell migration during Drosophila oogenesis Anna A. Ogienko*, Dmitry A. Karagodin, Valentina V. Lashina, Sergey I. Baiborodin, Eugeniya S. Omelina, Elina M. Baricheva CELL BIOL INT, 2013, Feb;37(2):149-59. doi: 10.1002/cbin.10025. |
|
| Identification of functionally significant elements in the second intron of the Drosophila melanogaster Trithorax-like gene D.A. Karagodin, E.S. Omelina, E.V. Fedorova, E.M. Baricheva GENE, 2013, Volume 520, Issue 2, Pages 178–184, doi: 10.1016/j.gene.2013.02.012 |
|
| Изменчивость нуклеотидной последовательности митохондриального гена субъединицы I цитохромоксидазы С у видов родов Aedes И Ochlerotatus (DIPTERA: CULICIDAE) Н.В. Храброва, Ю.В. Андреева, О.В. Ваулин, С.С. Алексеева, А.К. Сибатаев Vavilov journal of genetics and breeding, 2013, № 1, Т. 17. С 114-122. |
|
| Применение физических методов фармации для улучшения свойств лекарсвенных веществ Зевак Е.Г., Огиенко А.Г., Мызь С.А., Юношев А.С., Колесов Б.А., Огиенко А.А., Ачкасов А.Ф., Ильдяков А.В., Шинкоренко П.М., Красников А.А., Кучумов Б.М., Анчаров А.И., Манаков А.Ю., Болдырева Е.В. Химическая технология, 2013, Т. 14, №8. С. 480-492 |
|
| Parental Effects of Conditional Mutations and Their Explanations Chadov B.F., N.B. Fedorova, E.V. Chadova. RUSS J GENET+, 2013, Vol. 49, No. 2, pp. 141–150. © Pleiades Publishing, Inc. |
|
| Циклическая протомодель и феномен эволюции Чадов Б.Ф. Электронный журнал «Biocosmology- neo-Aristotelism, 2013, Vol. 3, No1, С.120-146. Режим доступа: http://www.biocosmology.ru |
|
| Циклическая модель образования материи и её актуальные приложения Чадов Б.Ф. Биосфера, 2013, Т.5, №3. Приложение. С.16. |
|
| Идея Творца в религии и науке Чадов Б.Ф. Эко-потенциал, 2013, Т.1. №1-2. С. 145 - 166 |
|
| Родительские эффекты условных мутаций у Drosophila melanogaster Чадов Б.Ф., Федорова Н.Б., Чадова Е.В. RUSS J GENET+, 2013, №2, том 49, с. 164-174 |
|
| ФИЛОГЕНЕТИЧЕСКИЕ ОТНОШЕНИЯ НЕКОТОРЫХ ВИДОВ РОДА Aquilegia СЕВЕРНОЙ АЗИИ ПО РАЗЛИЧНЫМ ДНК-МАРКЕРАМ Эрст А.С., Ваулин О.В. Vavilov journal of genetics and breeding, 2013, № 3. Т. 17. 477-486 с. |
|
| Дизайн нанокомпозитов сальбутамол/глицин для ингаляционной терапии: лекарственная форма нового поколения без использования хладонов Зевак Е.Г., Огиенко А.Г., Болдырева Е.В., Мызь С.А., Огиенко А.А., Коваленко Ю.Е.,Колесов Б.А., Дребущак В.А., Трофимов Н.А., Манаков А.Ю., Болдырев В.В. Нанотехнологии и охрана здоровья, 2013, №3, с.30-43. |
|
| Salbutamol-Glycine Composite Microballs for Pulmonary Drug Delivery Zevak E.G., Ogienko A.G., Boldyreva E. V., Myz S. A., Ogienko A.A., Kovalenko Y.E.,Kolesov B. A., Drebushchak V. A., Trofimov N.A., Krasnikov A. A., Manakov A. Y., Boldyrev V.V. Respiratory Drug Delivery Europe, 2013, V. 2. P. 329-334 |
|
| 2012 | Основные компоненты генной сети, контролирующей развитие дорзальных выростов хориона яиц Drosophila melanogaster Омелина Е.С., Баричева Э.М. RUSS J DEV BIOL+, 2012, Т. 43, № 3, С. 1-11 |
| Основные типы строения респираторных систем яйцевых оболочек насекомых и гены, участвующие в их формировании Омелина Е.С., Баричева Э.М., Федорова Е.В. Журнал общей биологии, 2012, Т. 73, № 3, С. 210-221 |
|
| Design of new drug forms by cryo-nanotechnologies A G Ogienko, E V Boldyreva, V V Boldyrev, A Y Manakov, S A Myz, A A Ogienko, E G Zevak and A S Yunoshev JPCS, 2012, 345 012044 doi:10.1088/1742-6596/345/1/012044 |
|
| Получение высокодисперсных форм лекарственных препаратов с использованием сублимационной сушки замороженных растворов в системах с клатратообразованием А. Г. Огиенко, Е. В. Болдырева, А. Ю. Манаков, С. А. Мызь, А. А. Огиенко, А. С. Юношев, Е. Г. Зевак, Н. В. Кутаев, А. А. Красников DOKL PHYS CHEM, 2012, том 444, № 5, с. 514–518 |
|
| Genetics of Intraspecific Similarity (Research Topic). Chadov B.F. Frontiers in Genetics, 2012, http://www.frontiersin.org/Journal/.2012 |
|
| Цикл и категории материалистической биокосмологии Чадов Б.Ф. Электронный журнал «Biocosmology- neo-Aristotelism, 2012, Vol. 2, No 1-2, P.51- 83. http://www.biocosmology.ru/. |
|
| Оценка времени жизни человечества по скорости вымирания языков Иванов Юрий Николаевич Вестник НО ПАНИ, 2012, № 15, с. 74 - 75 |
|
| Архитектура клеточного скелета Огиенко А.А., Баричева Э.М., Байбородин С.И. Наука из первых рук, 2012, 3(45), 100-103 |
|
| В ингаляторе - "нано" Огиенко А.Г., Зевак Е.Г., Огиенко А.А., Мызь С.А. Наука из первых рук, 2012, 3(45) 108-111 |
|
| A revision of chromosome II (CD) mapping in Chironomus plumosus (Linnaeus, 1758) group (Diptera, Chironomidae) Golygina V.V., Kiknadze I.I. COMP CYTOGENET, 2012, 6 (3): 249-266 |
|
| The karyotype and chromosome polymorphism of the Holarctic Species Chironomus “annularius” sensu Strenzke, 1959 (Diptera, Chironomidae) Kiknadze I.I., Istomina A.G., and Golygina V.V. Евразиатский энтомологический журнал, 2012, Т. 11 вып. 2: 95-114 |
|
| Полиморфизм и межвидовая изменчивость нуклеотидной последовательности гена субъединицы I цитохромоксидазы (COI) у видов двойников A и B Anopheles messeae и An. beklemishevi (Diptera: Culiucidae) Ваулин О.В., Новиков Ю.М. Vavilov journal of genetics and breeding, 2012, № 2. Т. 16. С. 358-368. |
|
| О концепциях биогенеза Иванов Юрий Николаевич Вестник НО ПАНИ, 2012, № 15, с. 57 - 62 |
|
| Coat color mutations and the monoamine content in the brain of farm-bred mink (Neovison vison) Trapezov O.V., Trapezova L.I., Alekhina T.A., Klochkov D.V., Ivanov Yu.N. Scientifur, 2012, No 3, V. 36 (3/4), P. 294-299. |
|
| The Biocosmology Categories Chadov Электронный журнал «Biocosmology- neo-Aristotelism, 2012, Vol. 2, No 3, P. 256-266. Режим доступа: http://www.biocosmology.ru/. |
|
| Циклическая модель образования материи и феномен сознания Чадов Б.Ф. Электронный журнал «Biocosmology- neo-Aristotelism, 2012, Vol. 2, No 4, P. 376 - 397. Режим доступа: http://www.biocosmology.ru/. |
|
| Цикл и категории материалистической биокосмологии Чадов Б.Ф. Академия Тринитаризма, 2012, публ. 17743, 17.11.2012. Эл № 77- 6567 http://www.trinitas.ru/rus/000/a0000001.htm |
|
| 2011 | Analysis and recognition of the GAGA transcription factor binding sites in Drosophila genes Omelina E.S., Baricheva E.M., Oshchepkov D.Yu., Merkulova T.I. COMPUT BIOL CHEM, 2011, V. 35, № 6, P. 363-370 |
| Для формирования дорзальных выростов хориона Drosophila melanogaster требуется белок GAGA Омелина Е.С., Павлова Н.В., Огиенко А.А., Баричева Э.М. Doklady Biochemistry and Biophysics, 2011, Т. 436, № 5, С. 696-698 |
|
| A new method of producing monoclinic paracetamol suitable for direct compression Ogienko А.G., Boldyreva Е.V., Manakov А.Yu., Boldyrev V.V., Yunoshev А.S., Ogienko А.А., Myz S.A., Ancharov А.I., Achkasov A.F., Drebushchak T.N. PHARM RES-DORDR, 2011, Volume 28, Number 12, Pages 3116-3127 |
|
| Hedvall effect’ in cryogrinding of molecular crystals. A case study of a polymorphic transition in chlorpropamide Drebushchak T.N., Ogienko A.A., Boldyreva E.V. CRYSTENGCOMM, 2011, V.13 (13), p. 4405-4410 |
|
| Новый способ получения высокодисперсного парацетамола для прессования без наполнителей А.Г. Огиенко, Е.В. Болдырева, А.Ю. Манаков, В.В. Болдырев, М.А. Михайленко, А.С. Юношев, А.А. Огиенко, А.И. Анчаров, А.Ф. Ачкасов, А.В. Ильдяков, А.А. Бурдин, Н.А. Туманов, А.С. Стопорев, Н.В. Кутаев DOKL PHYS CHEM, 2011, том 437, № 6, стр. 785-788 |
|
| An Integrated Chromosome Map of Microsatellite Markers and Inversion Breakpoints for an Asian Malaria Mosquito, Anopheles stephensi MARYAM KAMALI, MARIA V. SHARAKHOVA, ELINA BARICHEVA, DMITRII KARAGODIN, ZHIJIAN TU, IGOR V. SHARAKHOV J HERED, 2011, Vol. 102, № 6, P. 719-726 |
|
| Conditional Mutations in Drosophila B.F. Chadov, N.B. Fedorova, E.V. Chadova, H.A. Khotskina Journal of Life Sciences, 2011, Vol.5, № 3, March 2011, P. 224-239 |
|
| На пути к «естественной» философии Чадов Борис Федорович Электронный журнал «Biocosmology- neo-Aristotelism, 2011, Vol.1, No 2/3, P.221-273. |
|
| Медицина и биокосмология (отзыв на книгу В.К.Козлова и С.В.Ярилова "Введение в системную медицину") Чадов Б.Ф. Электронный журнал «Biocosmology- neo-Aristotelism, 2011, Т.1. №4, С.475-481 |
|
| 2010 | Локализация и частоты инверсионных разрывов в геномах видов Chironomus (Diptera, Chironomidae) Брошков А.Д., Мирошниченко Л.А., Кикнадзе И.И., Гусев В.Д. Информационный вестник ВОГИС, 2010, 14, 4: 654-662 |
| Особенности строения центромерных районов политенных хромосом хирономид (Diptera, Chironomidae) Голыгина В.В., Кикнадзе И.И., Брошков А.Д., Карамышева Т.В., Рубцов Н.Б. Информационный вестник ВОГИС, 2010, 14(4): 622 – 631 |
|
| Анализ структуры глаза мутантов по гену trithorax-like Drosophila Melanogaster Н.В. Павлова, Д.А. Карагодин, С.И. Байбородин, Э.М. Баричева Информационный вестник ВОГИС, 2010, Том 14, № 3, C. 558-568 |
|
| Числа хромосом некоторых видов семейства Ranunculaceae Juss. из Сибири. Эрст А. С., Брошков А. Д. Turczaninowia, 2010, Т. 13., Вып. 3., С. 152-153. |
|
| Географическая изменчивость ITS2 рДНК и COI мтДНК и криптические виды малярийного комара Anopheles messae Fall. (Diptera: Culicidae) Ваулин О.В., Новиков Ю.М. Информационный вестник ВОГИС, 2010, Т. 14. № 3. С. 546-557. |
|
| ДНК-диагностика микст-инвазий Opisthorchis felineus и Metorchis bilis с помощью метода ПЦР Брусенцов И.И., Катохин А.В., Сахаровская З.В., Сазонов А.Э., Огородова Л.М., Федорова О.С., Колчанов Н.А., Мордвинов В.А. Медицинская паразитология и паразитарные болезни, 2010, №2. С. 10-13. |
|
| Изменение энергетического статуса дрозофилы в результате генетической мутации Чадов Б. Ф., Федорова Н. Б., Чадова Е. В., Хоцкина Е. А., Мошкин М. П., Петровский Д. В. RUSS J GENET+, 2010, том 46, № 9, с. 1196–1201 |
|
| 2009 | Кариотип и хромосомный полиморфизм у хирономиды Chironomus melanescens Keyl, 1961 (Diptera, Chironomidae) Кикнадзе И.И., Брошков А.Д. Евразиатский энтомологический журнал, 2009, Т. 8. Прилож. 1: 19-28. |
| Поддержание паттернов экспрессии гомеозисных генов в развитии Drosophila melanogaster белками групп Polycomb, trithorax и ETP Фёдорова Е. В., Пиндюрин А. В., Баричева Э. М. RUSS J GENET+, 2009, Т. 45, № 10, С. 1301-1318. |
|
| Условно летальные мутации переводят геном из стабильного состояния в нестабильное Чадов Б.Ф., Чадова Е.В., Хоцкина Е.А., Федорова Н.Б. RUSS J GENET+, 2009, №3,Т. 45, 318-329 |
|
| Энергетическое предназначение живого и генетическая система Чадов Б.Ф. Вестник ЛНУ им. В. Даля, 2009, №1, С.72-105 |
|
| Влияние генотипического окружения хозяина - Drosophila melanogaster - на биологические эффекты эндосимбионта Wolbachia (штамм wMelPop) Воронин Д.А., Бочериков А.М., Баричева Э.М., Захаров И.К., Киселёва Е.В. Cell and Tissue Biology, 2009, Т. 51. № 4. С. 335-345. |
|
| Дифференциация кариофондов и геномной ДНК в природных популяциях Chironomus entis Shobanov (Diptera, Chironomidae) Голарктики Гундерина Л.И., Кикнадзе И.И., Голыгина В.В., Батлер М.Д. RUSS J GENET+, 2009, Т. 45. № 12., С. 1600-1608. |
|
| 2008 | Источники генетической изменчивости в природных популяциях Drosophila melanogaster Захаров И.К., Ваулин О.В., Илинский Ю.Ю., Синянский Я.Я., Коромыслов Ю.А., Коваленко Л.В., Иванников А.В., Захаренко Л.П., Волошина М.А., Чересиз С.В., Юрченко Н.Н. Информационный вестник ВОГИС, 2008, № 1/2. Т. 12. С. 112-126. |
| Молекулярно-генетическая характеристика новой гипоморфной мутации гена Trithorax-like и анализ ее влияния на оогенез Drosophila melanogaster Огиенко А.А., Карагодин Д.А., Павлова Н.В., Фёдорова С.А., Волошина М.А., Баричева Э.М. RUSS J DEV BIOL+, 2008, Т. 39. № 2. С. 134-142. |
|
| Системная регуляция генетических и цитогенетических процессов сигнальным каскадом ремоделирования актина: локус agnostic дрозофилы Медведева А. В., Молотков Д. А., Никитина Е. А., Попов А. В., Карагодин Д. А., Баричева Е. М., Савватеева-Попова Е. В. RUSS J GENET+, 2008, №6, том 44, с. 771-783. |
|
| Структура кариотипа хирономиды Chironomus uliginosus Keyl, 1960 (Diptera, Chironomidae) Брошков А.Д., Кикнадзе И.И., Истомина А.Г., Гундерина Л.И. Евразиатский энтомологический журнал, 2008, Т. 7, вып. 1: 57-65 |
|
| Interaction between the Drosophila heterochromatin proteins SUUR and HP1 Pindyurin A.V., Boldyreva L.V., Shloma V.V., Kolesnikova T.D., Pokholkova G.V., Andreyeva E.N., Kozhevnikova E.N., Ivanoschuk I.G., Zarutskaya E.A., Demakov S.A., Gorchakov A.A., Belyaeva E.S., Zhimulev I. F. J CELL SCI, 2008, 121, 1693-1703 |
|
| Mystery of Chironomus dorsalis Meigen karyotype (Diptera: Chironomidae) Kiknadze I.I., Golygina V.V., Broshkov A.D., Gunderina L.I., Istomina A.G. COMP CYTOGENET, 2008, № 1, V. 2, p 21-35 |
|
| Нарушение функционирования соматических клеток в яйцевых камерах Drosophila melanogaster у мутантов по гену trithorax-like Огиенко А.А., Лаухина О.В., Васильев Г.В., Баричева Э.М. Информационный вестник ВОГИС, 2008, т.12, с. 399-405 |
|
| The revision of chromosome I (AB) mapping in Chironomus plumosus group (Diptera, Chironomidae) Golygina V.V., Kiknadze I.I. COMP CYTOGENET, 2008, V. 2. №1: 37-55. |
|
| Цикличность живого и сущего Чадов Б.Ф. Философия науки, 2008, №2 (37). С.134-161 |
|
| Временнáя динамика и изменчивость по мультилокусным ДНК-маркерам ISSR-PCR в популяции Drosophila melanogaster г. Умань на протяжении двух десятилетий (1984-2004 гг.) Ваулин О.В., Захаров И.К. RUSS J GENET+, 2008, № 3. Т. 44. С. 359-365. |
|
| Ген SuUR - уникальный инструмент для изучения структуры и организации хромосом и генома дрозофилы Жимулёв И.Ф., Беляева Е.С., Андреенкова Н.Г., Андреева Е.Н., Белякин С.Н., Болдырева Л.В., Брусенцова И. В., Волкова Е.И., Демаков С.А., Демакова О.В., Заруцкая Е.А., Зыков И. А., Кокоза Е.Б., Колесникова Т.Д., Комор У.А., Коряков Д.Е., Макунин И.В., Пинд Информационный вестник ВОГИС, 2008, Т.12, N 1/2, c. 127-149 |
|
| Географическая изменчивость последовательностей дисков политенных хромосом комара-звонца Chironomus pseudothummi Str. (Diptera, Chironomidae) Кикнадзе И.И., Брошков А.Д., Истомина А.Г., Гундерина Л.И., Валлендук Х. Cell and Tissue Biology, 2008, Т. 50. № 6: 539-548 |
|
| 2007 | Основные аспекты развития половой системы самок Drosophila melanogaster Огиенко А.А., Федорова С.А., Баричева Э.М RUSS J GENET+, 2007, т. 43.№10. С.1341-1357 |
| Полиморфизм и дифференциация мультилокусных маркёров ДНК в природных популяциях Drosophila melanogaster Ваулин О.В., Гундерина Л.И., Захаров И.К. RUSS J GENET+, 2007, № 1, Т. 43, стр. 61-69 |
|
| Cytogenetic divergence of genomes in Chironomus plumosus group (Diptera: Chironomidae) Golygina V.V., Kiknadze I.I., Istomina A.G., Gunderina L.I., Miroshnichenko L.A., Gusev V.D. COMP CYTOGENET, 2007, V. 1, №1, P. 17-32 |
|
| High-resolution analysis of Drosophila heterochromatin organization using Su(UR Su(var)3-9 double mutants EN Andreyeva, TD Kolesnikova, OV Demakova, M Mendez-Lago, GV Pokholkova, ES Belyaeva, F Rossi, N Corradini, P Dimitri, A Villasante, IF Zhimulev. P NATL ACAD SCI USA, 2007, 104: 12819-12824 |
|
| SUUR joins separate subsets of PcG, HP1 and B-type lamin targets in Drosophila Pindyurin A.V., Moorman C., de Wit E., Belyakin S.N., Belyaeva E.S., Christophides G.K., Kafatos F.C., van Steensel B., Zhimulev I.F. J CELL SCI, 2007, 120, 2344-2351 |
|
| The structural peculiarities of karyotypes in species of Propsilocerus akamisi sibling group (Diptera: Chironomidae) Kiknadze I.I., Istomina A.G., Golygina V.V., Rubtsov N.B., Karamysheva T.V. COMP CYTOGENET, 2007, V. 1, №1, P. 33-43 |
|
| The SU(VAR)3-9/HP1 complex differentially regulates the compaction state and degree of underreplication of X chromosome pericentric heterochromatin in Drosophila melanogaster OV Demakova, GV Pokholkova, TD Kolesnikova, SA Demakov, EN Andreyeva, ES Belyaeva, IF Zhimulev GENETICS, 2007, 175: 609-620 |
|
| Квазицикл «ген-проген» – имманентное свойство живого Чадов Б.Ф. Философия науки, 2007, № 1 (32), С.129-156 |
|
| Временнáя изменчивость геномной ДНК Drosophila melanogaster в популяции г. Умань (Украина) Ваулин О.В., Гундерина Л.И., Захаров И.К. Вестник ТГУ, 2007, № 300 (2).(июль). С. 107-108. |
|
| Изменчивость гена Adh в природных популяциях Drosophila melanogaster Ваулин О.В., Захаров И.К. Вестник ТГУ, 2007, № 300 (2).(июль) С. 109-112. |
|
| 2006 | Полученbе и анализ новых мутаций по гену Trithorax-like Drosophila melanogaster Федорова Е.В., Огиенко А.А., Карагодин Д.А., Айманова К.Г., Баричева Э.М. RUSS J GENET+, 2006, №2, т. 42, с. 149-158 |
| Хромосомный полиморфизм и дивергенция популяций у Chironomus nuditarsis Str. (Diptera, Chironomidae) Кикнадзе И.И., Михайлова П., Истомина А.Г., Голыгина В.В., Инт Панис Л., Крастанов Б. Cell and Tissue Biology, 2006, Т. 48. № 7. С. 595-609. |
|
| Эффект положения мозаичного типа brownD и его связь с ассоциациями гетерохроматиновых районов у Drosophila melanogaster Е.С. Беляева, Г.В. Похолкова, И.Ф. Жимулев Doklady Akademii Nauk, 2006, Т. 409, N 4, 559-562 |
|
| DNA underreplication in intercalary heterochromatin regions in polytene chromosomes of Drosophila melanogaster correlates with the formation of partial chromosomal aberrations and ectopic pairing Belyaeva ES, Demakov SA, Pokholkova GV, Alekseyenko AA, Kolesnikova TD, Zhimulev IF CHROMOSOME RES, 2006, 115(5):355-66 |
|
| Анализ новой гипомофной мутации гена Trithorax-like, влияющей на оогенез Drosophila melanogaster Огиенко А.А., Карагодин Д.А., Федорова С. А., Федорова Е. В., Лашина В. В., Баричева Э. М. RUSS J DEV BIOL+, 2006, № 3, т. 37, с. 157–166 |
|
| Variability and differentiation of genomic DNA in the Drosophila melanogaster populations of Russia and Ukraine. Vaulin O.V., Zharikov T.YU., Gunderina L.I., Zakharov I.K. Dros Inf Serv, 2006, 2006, V. 89. p. 59-62. |
|
| Новый этап в развитии генетики и термин “эпигенетика” Чадов Борис Фёдорович RUSS J GENET+, 2006, . Т.42. №9. С. 1261-1275 |
|
| Nucleotide sequence variability of Adh gene of Drosophila melanogaster in the populations of Eurasia Vaulin O.V., Zakharov I.K. Dros Inf Serv, 2006, No. 89. P. 51-54. |
|
| Анализ новой гипомофной мутации гена Trithorax-like, влияющей на оогенез Drosophila melanogaster Огиенко А.А., Карагодин Д.А., Федорова С. А., Федорова Е. В., Лашина В. В., Баричева Э. М. RUSS J DEV BIOL+, 2006, № 3, т. 37, с. 157–166 |
|
| Ген SuUR и его участие в организации эпигенетически репрессированных районов хромосом Drosophila melanogaster Колесникова Т.Д., Андреева Е.Н., Пиндюрин А.В., Ананько Н.Г., Белякин С.Н., Шлома В.В., Юрлова А.А., Макунин И.В., Похолкова Г.В., Волкова Е.И., Заруцкая Е.А., Кокоза Е.Б., Семешин В.Ф., Беляева Е.С., Жимулев И.Ф. RUSS J GENET+, 2006, 42 (8): 1013–1028 |
|
| 2005 | GAGA фактор, кодируемый геном Trithorax-like, требуется для оогенеза дрозофилы Огиенко А.А., Карагодин Д.А., Федорова С.А., Лашина В.В., Федорова Е.В., Киселева Е.В., Губанова Н.В., Баричева Э.М. RUSS J DEV BIOL+, 2005, № 3 ,том 36, стр.388 |
 Монографии
Монографии
| 2025 | Генетика внутривидового сходства (опыты с условными мутациями у дрозофилы) Чадов Борис Федорович |
| 2024 | Genes and Ontogenes in Drosophila: Exploring the Role of RNA Forms Nina B. Fedorova a*, Eugenia V. Chadova a and Boris F. Chadov a |
| Ontogenes and Their Role in Morphogenesis Chadov B. F., Fedorova N. B. |
|
| 2022 | Conditional Mutations and New Genes in Drosophila Boris F. Chadov, Nina B. Fedorova |
| 2020 | Inbreeding Depression and Heterosis as
the Phenomena of Two-Component Genome B. F. Chadov and N. B. Fedorova |
| 2016 | Karyotypes of Palearctic and Holarctic species of the genus Chironomus [Electronic resource] Кикнадзе И.И., Истомина А.Г.. Голыгина В.В.. Гундерина Л.И. |
| 2015 | Мысли о науке и жизни, 8-е издание, дополненное Иванов Юрий Николаевич |
| Мутационный процесс и экологические аспекты генетики Иванов Юрий Николаевич |
|
| Циклическая протомодель и феномен эволюции Чадов Борис Федорович |
|
| 2014 | Мысли о науке и жизни, 7-е издание, дополненное Иванов Юрий Николаевич |
| 2012 | Мысли о науке и жизни Иванов Юрий Николаевич |
| Эпигенетическая феноменология у условных мутантов Drosophila melanogaster: морфозы и модификации Чадов Б.Ф., Чадова Е.В., Федорова Н.Б. |
|
| Эпигенетическая феноменология у условных мутантов Drosophila melanogaster: морфозы и модификации Чадов Б. Ф., Чадова Е. В., Федорова Н. Б. |
|
| 2011 | Полиморфизм природных популяций комаров (Chironomus и Anopheles). Голыгина В.В., Ваулин О.В., Гундерина Л.И., Истомина А.Г., Брошков А.Д., Новиков Ю.М., Кикнадзе И.И., Захаров И.К. |
| Полиморфизм природных популяций Drosophila melanogaster – влияние факторов внешней среды, взаимодействие генов и геномов, роль мобильных генетических элементов Илинский Ю.Ю., Коромыслов Ю.А., Ваулин О.В., Антоненко О.В., Иванников А.В., Юрченко Н.Н., Захаренко Л.П., Васильева Л.А., Захаров И.К. |
|
| Микроскопический анализ биологических объектов. Научные результаты Нарушение формирования актинового цитоскелета в питающих клетках TRL мутантов Drosophila melanogaster Огиенко А.А., Лашина В.В., Байбородин С.И., Баричева Э.М. |
|
| 2010 | Chapter 4 "Cytoplasmic Transport during Drosophila Oogenesis" Fedorova S.A., Karagodin D.A., Ogienko A.A., Baricheva E.M. |
| Глава. Квазицикл «ген-проген» и эволюция Чадов Борис Федорович |
|
| 2009 | Популяционно-генетический анализ: методы электрофореза белков и полимеразной цепной реакции в популяционной генетике. Учебно-методическое пособие для студентов биологического факультета. // Отв. ред.: И.В. Пак, И.К. Захаров / Тюмень: Изд-во Тюменского государственного университета. 2009. 69 с. Жигилева О.Н., Пак И.В., Ваулин О.В. |
| 2008 | Karyotype structure in species of genus Propsilocerus. (Diptera, Chironomidae) Kiknadze I.I., Istomina A.G., Wang X., Golygina V.V. |
| Divergence of centromeric heterochromatin is an important way of evolution in the genus Propsilocerus (Diptera, Chironomidae) Golygina V.V., Rubzov N.B., Karamisheva T.V., Kiknadze I.I. |
|
| Chironomus sp. K – a new member of plumosus sibling-group from Japan Golygina V.V., Ueno R. |
|
| 2006 | Цитогенетические механизмы дивергенции популяций и видов у хирономид (Diptera: Chironomidae) Кикнадзе И.И., Гундерина Л.И., Истомина А.Г., Голыгина В.В., Андреева Е.Н., Гусев В.Д., Мирошниченко Л.А. |
| Chironomidae: виды, популяции, генетическая изменчивость Голыгина В.В., Истомина А.Г. |
|
| Исследования взаимодействий генов Drosophila melanogaster Волошина М.А., Огиенко А.А., Баричева Э.М. |
Конференции
| 2022 | Transcriptome analysis of Trl mutants reveals the role of mitochondria in the mass death of germ cells Fedorova S., Dorogova N., Karagodin D., Baricheva E. Bioinformatics of Genome Regulation and Structure/Systems Biology (BGRS/SB-2022) |
| 2021 | Функциональный анализ однонуклеотидных замен rs2072580 и rs590352,
связанных с предрасположенностью к развитию онкологических заболеваний Дегтярева А.О., Леберфарб Е.Ю., Брусенцов И.И., Антонцева Е.В., Меркулова Т.И. NGS в медицинской генетике. Пятая международная научно-практическая конференция. |
| 2020 | Functional study of potential regulatory SNPs (rs590352, rs11542583, rs3829202, rs78317230, rs2072580, rs4796672) Arina Degtyareva, Elena Leberfarb, Ilya Brusentsov, Tatiana Kuzina, Tatiana Merkulova 2020 Cognitive Sciences, Genomics and Bioinformatics, CSGB 2020 |
| 2019 | ПОЛИМОРФИЗМ rs590352 (C>G) ГЕНА ATXN7L3B И РИСК РАЗВИТИЯ КОЛОРЕКТАЛЬНОГО РАКА Дегтярева А.О., Леберфарб Е.Ю., Брызгалов Л.О., Брусенцов И.И., Воевода М. И., Аутеншлюс А. И., Максимов В. Н., Морозов Д. В., Соколов А.В., Меркулова Т.И. ВОГИС |
| Функциональная аннотация регуляторных полиморфизмов, связанных с развитием онкологических заболеваний. Леберфарб Е.Ю., Дегтярева А.О., Брусенцов И.И., Ефимова Е.Г., Меркулова Т.И. VI Съезд биофизиков России |
|
| Hi-C analysis and physical mapping identify inversions in the Aedes aegypti genome. Atashi Sharma Varvara Lukyanchikova Ilja Brusentsov Igor V. Sharakhov Maria V. Sharakhova Annual Meeting of the Eastern Branch of Entomological Society of America |
|
| 2018 | Germ cell migration under GAGA-factor control A. Khruscheva, K. Ahmetova , S. Fedorova, E. Baricheva , N. Dorogova 11-ая Международная конференция по биоинформатике регуляции и структуры геномов и системной биологии БГРС 2018, |
| Зиготический отбор у Drosophila melanogaster и новая редакция дарвиновской концепции видообразования Чадов Б.Ф., Федорова Н.Б. Эволюция жизни на Земле |
|
| Зиготический отбор у Drosophila melanogaster и новая редакция дарвиновской концепции видообразования Б.Ф.Чадов, Н.Б.Федорова V Международный симпозиум Эволюция жизни на Земле |
|
| "Влияние соматического окружения на миграцию предшественников зародышевых клеток в эмбриогенезе Drosophila melanogaster" Ахметова К.А., Хрущева А.С., Дорогова Н.В., Баричева Э.М., Фёдорова С.А 4-ая Международная конференция "Хромосома 2018" |
|
| " Хроматин-ремоделирующий фактор GAGA регулирует различные типы миграции при формировании половых клеток самок дрозофилы " Галимова Ю.А., Хрущева А.С., Дорогова Н.В ., Огиенко А.А., Фёдорова С.А. 4-ая Международная конференция "Хромосома 2018" |
|
| 2017 | NOVEL APPROACH TO FUNCTIONAL SNPs DISCOVERY FROM GENOME-WIDE DATA REVEALS PROMISING VARIANTS FOR COLON CANCER RISK E.E. Korbolina I.I. Brusentsov L.O. Bryzgalov E.Yu. Leberfarb A.O. Degtyareva T.I. Merkulova BELYAEV CONFERENCE |
| Gene localization in genomes of species from the genus Chironomus (Diptera, Chironomidae) Veronika Golygina, Larisa Gunderina, Oksana Ermolaeva, Andrey Broshkov1 20th International Symposium on Chironomidae |
|
| Analysis of gene localization in genoms of species from the genus Glyptotendipes (Diptera, Chironominae) Oksana V. Ermolaeva, Larisa I. Gunderina, Veronika V. Golygina 20th International Symposium on Chironomidae |
|
| Сравнительный анализ локализации генов в кариотипах видов трибы Chironomini (Diptera, Chironomidae) О.В. Ермолаева, Л.И. Гундерина, В.В. Голыгина Материалы XV съезда Русского энтомологического общества |
|
| Мутации гена trl, кодирующего GAGA-фактор дрозофилы, вызывают преждевременную миграцию клеток зародышевого пути Хрущева А.С. Дорогова Н.В. Баричева Э.М. Международная конференция «Беляевские чтения», посвященная 100-летию академика АН СССР Д.К. Беляева |
|
| Пространственная и временная динамика соотношений численности видов малярийных комаров в окрестностях г. Новосибирска О.В. Ваулин, И.К. Захаров БЕЛЯЕВСКИЕ ЧТЕНИЯ Международная конференция, посвященная 100-летию со дня рождения академика АН СССР Д.К. Беляева |
|
| Повышенная мутабильность в линии Drosophila melanogaster 3NS и ее модификация при межлинейных скрещиваниях Ю.А. Коромыслов, О.В. Ваулин, И.К. Захаров БЕЛЯЕВСКИЕ ЧТЕНИЯ, Международная конференция, посвященная 100-летию со дня рождения академика АН СССР Дмитрия Константиновича Беляева |
|
| Ортогенез и дарвинизм: перспектива синтеза в свете данных по условным мутациям Чадов Б.Ф., Чадова Е.В., Федорова Н.Б. Всероссийская (с междунар. участием) научная конференция Современные проблемы эволюции и экологии, ХХХ Любищевские чтения |
|
| 2016 | Slbo-gal4 driver in line 6458: a cautionary note on its use in Drosophila oogenesis Огиенко АА, Яринич Л.А., Федорова Е.В., Лебедев М.О., Пиндюрин А.В., Баричева Э.М. International mini-conference chromosomes and mitosis |
| Использование NGS методов выявления функционально активных районов генома для поиска полиморфизмов, определяющих развитие патологий, на примере колоректального рака Е.Ю. Леберфарб, Л.О. Брызгалов, И.И.Брусенцов, Ю.Л. Орлов, Т.И. Меркулова NGS в медицинской генетике |
|
| Эволюция как феномен образующейся Вселенной: стратегия и тактика эволюции живого Чадов Борис Федорович Международная научная конференция XXX Любищевские чтения |
|
| Genetics: the energy aspect Nina B. Fedorova, Boris F. Chadov Regional Interdisciplinary Conference – Humboldt Kolleg«Energy conversion: from nature to technology » |
|
| Transcriptomics: Genes and Ontogenes in Drosophila Nina B. Fedorova, Eugenia V. Chadova, and Boris F. Chadov 2nd International Conference on Transcriptomics |
|
| Эволюция хромосомной локализации генов в роде Chironomus Голыгина В.В., Гундерина Л.И., Брошков А.Д., Ермолаева О.В. VI международная конференция по кариосистематике беспозвоночных животных |
|
| Молекулярно-генетические нарушения в стабильных и нестабильных аллелях white-гена Drosophila melanogaster Ваулин О.В, Волошина М.А., Коромыслов Ю.А., Захаров И.К. Всероссийская конференция с международным участием «50 лет ВОГиС: успехи и перспективы» |
|
| Высокая мутабильность, возникающая при различных вариантах скрещиваний линии 3NS Drosophila melanogaster Коромыслов Ю.А., Ваулин О.В., Захаров И.К. Всероссийская конференция с международным участием «50 лет ВОГиС: успехи и перспективы» |
|
| 2015 | Выявление и функциональный анализ регуляторных SNPs, связанных с развитием рака толстого кишечника Леберфарб Е.Ю., Брызгалов Л.О., Брусенцов И.И., Дегтярева А.О., Меркулова Т.И. VII Съезд Российского общества медицинских генетиков |
| Исследование генетических систем, контролирующих движение бордюрных клеток в яичниках дрозофилы Огиенко А.А.,Федорова Е.В., Пиндюрин А.В., Яринич Л.А.,Баричева Э.М. Хромосома 2015 |
|
| Композиция бетулина с биосовместимыми носителями с улучшенной динамикой растворения. Шахтшнейдер Т.П., Мызь С.А., Михайленко М.А., Кузнецова С.А., Огиенко А.Г., Огиенко А.А., Болдырева Е.В., Болдырев В.В. Инновации в медицине: основные проблемы и пути их решения. Регенеративная медицина и новые биосовместимые материалы : материалы III Международного форума |
|
| Design salbutamol and budesonide solid dispersion for pulmonary drug delivery. Bogdanova E., Ogienko A., Boldyreva E., Myz S., Ogienko A., Kolesov B., Drebushchak V., Manakov A., Kovalenko Yu., Trofimov N., Boldyrev V. MedChem 2015 |
|
| Использование клатратных гидратов в вакуум-сублимационном способе получения ультрадисперсных порошков на примере аммиачной селитры Огиенко А.Г., Богданова Е.Г., Дребущак В.А., Ильдяков А.В., Юношев А.С., Огиенко А.А., Красников А.А., Манаков А.Ю. IV Всероссийская научно-практическая конференция молодых ученых и специалистов «Материалы и технологии XXI века» |
|
| Сравнение локализации генов, кодирующих рибосомные белки и рРНК, в хромосомах видов рода Chironomus. Гундерина Л.И.,Голыгина В.В., Брошков А.Д., Ермолаева О.В. Международная конференция «Хромосома 2015» |
|
| Ненаследование приобретенных признаков Иванов Юрий Николаевич XXIV МЕЖДУНАРОДНЫЙ СИМПОЗИУМ «Охрана био - ноосферы. Нетрадиционное растениеводство. Эниология. Экология и здоровье» |
|
| Менделевские и не-менделевские гены Иванов Юрий Николаевич XXIV МЕЖДУНАРОДНЫЙ СИМПОЗИУМ «Охрана био - ноосферы. Нетрадиционное растениеводство. Эниология. Экология и здоровье» |
|
| Генная инженерия как орудие геноцида Иванов Юрий Николаевич XXIV МЕЖДУНАРОДНЫЙ СИМПОЗИУМ «Охрана био - ноосферы. Нетрадиционное растениеводство. Эниология. Экология и здоровье» |
|
| 2014 | Цитогенетическая идентификация видов рода Chironomus (Diptera, Chironomidae) И. И. Кикнадзе, А. Г. Истомина, В. В. Голыгина, Л. И. Гундерина ХI съезд Гидробиологического общества при Российской академии наук |
| Chromosomal localization of ribosomal RNA genes in the genus Chironomus (Diptera, Chironomidae) Larisa I. Gunderina, Veronika V.Golygina 19th International Symposium on Chironomidae |
|
| Construction and comparison of gene maps of Chironomus riparius and Chironomus balatonicus (Diptera, Chironomidae) Veronika V.Golygina, Larisa L. Gunderina, Andrey D. Broshkov 19th International Symposium on Chironomidae |
|
| Karyotypes of Palearctic and Holarctic species from the genus Chironomus Iya I. Kiknadze, Albina G. Istomina, Veronika V. Golygina, Larisa I. Gunderina 19th International Symposium on Chironomidae |
|
| Высокоэффективные лекарственные формы нового поколения: фармацевтические композиции сальбутамола и будесонида для использования в порошковых ингаляторах. Огиенко А.Г., Богданова Е.Г., Болдырева Е.В., Мызь С.А., Трофимов Н.А., Огиенко А.А., Манаков А.Ю., Болдырев В.В. Инновации в медицине: основные проблемы и пути их решения. Регенеративная медицина и новые биосовместимые материалы : материалы III Международного форума |
|
| Общая теория видовых миграций Иванов Юрий Николаевич XXIII МЕЖДУНАРОДНЫЙ СИМПОЗИУМ «Охрана био - ноосферы. Нетрадиционное растениеводство. Эниология. Экология и здоровье» |
|
| Критика концепции малых мутаций Иванов Юрий Николаевич XXIII МЕЖДУНАРОДНЫЙ СИМПОЗИУМ «Охрана био - ноосферы. Нетрадиционное растениеводство. Эниология. Экология и здоровье» |
|
| Эволюция в свете циклической протомодели Чадов Б.Ф. Современные проблемы эволюции и экологии |
|
| A cyclic protomodel for the physical bases of morality Chadov B.F. The 8th International Seminar on Biocosmology |
|
| ДНК-маркеры и принципы реконструкции филогенетических отношений Ваулин Олег Викторович III (V) Всероссийская молодёжная конференция с участием иностранных учёных "Перспективы развития и проблемы современной ботаники" |
|
| 2013 | Циклическая модель образования материи и её актуальные приложения Чадов Б.Ф. III Международный научный конгресс «Глобалистика-2013» посвященный 150-летию со дня рождения Владимира Ивановича Вернадского (1863-1945). |
| Циклическая протомодель и феномен эволюции Тезисы докладов. С. 106 -108. Чадов Б.Ф. III Международный научный конгресс «Глобалистика-2013» посвященный 150-летию со дня рождения Владимира Ивановича Вернадского (1863-1945) |
|
| Циклическая модель образования материи и её актуальные приложения. Презентации Международного симпозиума «Мегаистория и глобальная эволюция». http://www.socionauki.ru/ Чадов Б.Ф. III Международного научного конгресса «Глобалистика-2013» посвященного 150-летию со дня рождения Владимира Ивановича Вернадского (1863-1945) |
|
| Cyclic protomodel and biocosmology. Round Table: Biocosmology – neo-Aristotelism. Organizer: Yoo Kwon Jong, Korea. . Chadov B.F. The 23rd World Congress of Philosophy |
|
| Оценка времени жизни человечества по убыли относительного прироста населения мира Иванов Юрий Николаевич XXII МЕЖДУНАРОДНЫЙ СИМПОЗИУМ «Охрана био - ноосферы. Нетрадиционное растениеводство. Эниология. Экология и здоровье» |
|
| Критика политипической концепции вида и возврат к линнеевской типологии в систематике Иванов Юрий Николаевич XXII МЕЖДУНАРОДНЫЙ СИМПОЗИУМ «Охрана био - ноосферы. Нетрадиционное растениеводство. Эниология. Экология и здоровье» |
|
| Симпатрическая дивергенция видов как мнимое явление Иванов Юрий Николаевич XXII МЕЖДУНАРОДНЫЙ СИМПОЗИУМ «Охрана био - ноосферы. Нетрадиционное растениеводство. Эниология. Экология и здоровье» |
|
| Исследование генетических систем, контролирующих динамику и реорганизацию актинового цитоскелета в ходе оогенеза Drosophila melanogaster Огиенко Анна Александровна, Омелина Евгения Сергеевна, Федорова Елена Владимировна, Баричева Элина Михайловна Конференция ВОГиС Проблемы Генетики и Селекции |
|
| «Дизайн комбинированных нанокомпозитов, не требующих использования фреонов в составе противоастматических лекарственных форм» А.Г. Огиенко, Е.Г. Зевак, Е.В. Болдырева, С.А. Мызь, А.А. Огиенко, А.В. Ильдяков, Ю.Е. Коваленко, Б.А. Колесов, В.А. Дребущак, Н.А. Трофимов, А.С. Юношев, А.Ю. Манаков, академик В.В. Болдырев «Технологии и оборудование химической, биотехнологической и пищевой промышленности» |
|
| «New Drug Forms Design with Employment of SEM» A.А. Ogienko, S. Myz, A.G. Ogienko, E. Boldyreva, V. Boldyrev 10th International Conference on Nanoscience & Nanotechnology, Салонники, Греция, 9-12 июля |
|
| 2012 | The Dynamics of the dissolution of the ultrafine ibuprofen in comparision with initial substance Myz S.A., Ogienko A.G., Shakhtshneider T.P., Boldyreva E.V., Manakov A.Yu., Boldyrev V.V., Ogienko A.A., Yunoshev A.S., Krasnikov A.A., Ildyakov A.V., Zevak E.G., Ancharov A.I. Fourteenth Annual Conference YUCOMAT 2012 |
| Microscopy in the design of new drug forms Ogienko A.A., Myz S.A., Boldyreva E.V. Fourteenth Annual Conference YUCOMAT 2012 |
|
| Взаимодействие гена Trithorax-like с генами, участвующими в формировании дорзальных выростов хориона яйца Drosophila melanogaster Омелина Е.С., Баричева Э.М., Лашина В.В. III Международная конференция "Дрозофила в экспериментальной генетике и биологии" |
|
| Применение СЭМ в работах по дизайну лекарственных форм нового поколения. Огиенко АА, Мызь С.А., Болдырева Е.В. XXXIV Российская конференция по электронной микроскопии |
|
| The Biocosmology Categories Chadov B.F. 3rd International Seminar on Biocosmology. June 30th- July1st , 2012, Witten/Herdecke, Germany. Oral report. Book of Abstracts. P.28. Режим доступа: http://www.biocosmology.ru/. |
|
| Ожидания и факты генетики популяций Иванов Юрий Николаевич 21 Международный симпозиум "Охрана био-ноосферы. Нетрадиционное растениеводство. Селекция. Эниология. Экология и здоровье" |
|
| В-хромосомы и программа исследования их действия в регуляции плотности популяции вида Иванов Юрий Николаевич 21 Международный симпозиум "Охрана био-ноосферы. Нетрадиционное растениеводство. Селекция. Эниология. Экология и здоровье" |
|
| Оценка времени жизни человечества по скорости вымирания языков Иванов Юрий Николаевич 21 Международный симпозиум "Охрана био-ноосферы. Нетрадиционное растениеводство. Селекция. Эниология. Экология и здоровье" |
|
| ЭВОЛЮЦИЯ ХРОМОСОМНОЙ ОРГАНИЗАЦИИ СЕМЕЙСТВА ГЕНОВ РИБОСОМНОЙ РНК (rRNA) В ТРИБЕ CHIRONOMINI Гундерина Л.И., Голыгина В.В., Брошков А.Д., Ермолаева О.В., Кикнадзе И.И. Хромосома 2012, Новосибирск, 2-7 сентября |
|
| РЕВИЗИЯ КАРТИРОВАНИЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДИСКОВ ПОЛИТЕННЫХ ХРОМОСОМ ВИДОВ ГРУППЫ C. PLUMOSUS (DIPTERA, CHIRONOMIDAE) Голыгина В.В., Кикнадзе И.И. Хромосома 2012, Новосибирск, 2-7 сентября |
|
| АНАЛИЗ ЛОКАЛИЗАЦИИ ЯДРЫШКОВЫХ ОРГАНИЗАТОРОВ В КАРИОТИПАХ ХИРОНОМИД (DIPTERA, CHIRONOMIDAE) МЕТОДОМ ФЛУОРЕСЦЕНТНОЙ IN SITU ГИБРИДИЗАЦИИ Ермолаева О.В., Голыгина В.В., Гундерина Л.И., Брошков А.Д., Кикнадзе И.И. XIV Съезд русского энтомологического общества |
|
| Design of new drug forms by cryo-nanotechnology Ogienko A.G., Boldyreva E.V., Manakov A.Yu., Yunoshev A.S., Ogienko A.A., Myz S.A., Zevak E.G., Ancharov A.I., Boldyrev V.V. Fourteenth Annual Conference YUCOMAT 2012 |
|
| ДИВЕРГЕНЦИЯ ИНВЕРСИОННЫХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ ДИСКОВ ПОЛИТЕННЫХ ХРОМОСОМ У ВИДОВ РОДА CHIRONOMUS (DIPTERA, CHIRONOMIDAE) ИЗ ЦИТОКОМПЛЕКСА «PSEUDOTHUMMI» Брошков А.Д., Гундерина Л.И., Кикнадзе И.И. XIV Съезд Русского Энтомологического Общества, |
|
| Кариотип и хромосомный полиморфизм голарктического вида Chironomus “annularius” sensu Strenzke, 1959 (Diptera, Chironomidae) Кикнадзе И.И., Истомина А.Г., Голыгина В.В. XIV съезд Российского энтомологического общества |
|
| Оценка времени жизни человечества по скорости вымирания языков Иванов Юрий Николаевич III Международн. симпозиум "Экологические проблемы животных и человека" |
|
| Условные мутации у D. melanogaster: особенности локализации и вторичного мутирования. Чадов Б.Ф., Федорова Н.Б., Чадова Е.В. International Conference Chromosoma - 2012. |
|
| Consciousness in the light of cyclical model of matter: mechanism and evolution. Proceedings of Chung-Ang University, P. 7-9 Сhadov B.F. International Seminar on Biocosmology & The 3rd International Conference on Comparative Studies of Mind |
|
| 2011 | Естественный отбор: способ производства новых видов или перманентное состояние материи? Чадов Борис Федорович ХХV Любищевские чтения. Современные проблемы эволюции. |
| New drug forms design using cryo-nanotechnologies A.G. Ogienko, S.A. Myz, A.A. Ogienko, A.S. Yunoshev, N.V. Kutaev, E.G. Zevak Четвертый Международныйконкурс научных работ молодых ученых в области нанотехнологий, проводимый в рамках IV-го Международного Форума по Нанотехнологиям |
|
| Дизайн фармацевтических субстанций с использованием сублимационной сушки замороженных растворов в системах с клатратообразованием А.Г. Огиенко, Е.В. Болдырева, В.В. Болдырев, А.Ю. Манаков, С.А. Мызь, А.А. Огиенко, А.С. Юношев, Е.Г. Зевак, Н.В. Кутаев Научная инновационная школа-конференция в рамках Международного молодежного инновационного форума «ИНТЕРРА-2011» |
|
| Micronization of ibuprofen by spray freeze-drying Ogienko A.G., Boldyreva E.V., Manakov A.Yu., Boldyrev V.V., Ogienko A.A., Myz S.A., Yunoshev A.S., Kutaev N.V., Krasnikov A.A., Ildyakov A.V. International Congress on Organic Chemistry |
|
| A new method of producing monoclinic paracetamol suitable for direct compression Ogienko А.G., Boldyreva Е.V., Manakov А.Yu., Boldyrev V.V., Yunoshev А.S., Ogienko А.А., Myz S.A., Ancharov А.I., Achkasov A.F., Drebushchak T.N. 2nd World Conference on Physico-Chemical Methods in Drug Discovery and Development |
|
| ‘Hedvall effect’ in cryogrinding of molecular crystals. A case study of a polymorphic transition in chlorpropamide A.G. Ogienko, E.V. Boldyreva, V.V. Boldyrev, A.Yu. Manakov, A.S. Yunoshev, M.A. Mikhailenko, A.A. Ogienko, A.I. Ancharov, N.V. Kutaev, A.F. Achkasov, A.S. Stoporev, N.A. Tumanov, A.V. Ildyakov, A.A. Burdin VII International Conference on Mechanochemistry and mechanical Alloying (INCOME 2011) |
|
| Исследование процессов, происходящих при отжиге замороженных растворов (стекол) в системах с клатратообразованием в условиях пониженного давления, и их использование для получения высокодисперсных форм лекарственных препаратов А.Г. Огиенко, Е.В. Болдырева, В.В. Болдырев, А.Ю. Манаков, А.С. Юношев, М.А. Михайленко, А.А. Огиенко, С.А. Мызь, Н.В. Кутаев Технологии и оборудование химической, биотехнологической и пищевой промышленности: материалы 4-й Всероссийской научно-практической конференции студентов, аспирантов и молодых ученых с международным участием |
|
| Model API composite balls for pharmaceutical application Ogienko A.G., Boldyreva E.V., Manakov A.Yu., Boldyrev V.V., Yunoshev A.S., Mikhailenko M.A., Ogienko A.A., Ancharov A.I., Kutaev N.V., Krasnikov A.A. Current topics in organic chemistry |
|
| Использование метода замороженных растворов для получения метастабильных полиморфных модификаций, сольватов, упаковочных комплексов молекулярных кристаллов и высокодисперсных образцов лекарственных препаратов А.Г. Огиенко, Е.В. Болдырева, А.Ю. Манаков, В.В. Болдырев, М.А. Михайленко, А.С. Юношев, В.А. Дребущак, С.А. Мызь, А.А. Огиенко, А.И. Анчаров, Б.А. Захаров, А.Ф. Ачкасов, А.В. Ильдяков, А.А. Бурдин, Н.А. Туманов, Н.В. Кутаев, С.В. Ильдяков IV Всероссийская Конференция по Наноматериалам НАНО-2011 |
|
| Получение высокодисперсного парацетамола методом сублимационной сушки замороженных растворов в системах с клатратообразованием Н.В. Кутаев, А.Г. Огиенко, Е.В. Болдырева, А.Ю. Манаков, В.В. Болдырев, М.А. Михайленко, А.С. Юношев, А.А. Огиенко, А.С. Стопорев, А.И. Анчаров, А.Ф. Ачкасов, А.В. Ильдяков, А.А. Бурдин, Н.А. Туманов, С.В. Ильдяков IV Всероссийская Конференция по Наноматериалам НАНО-2011 |
|
| Получение высокодисперсных форм лекарственных препаратов сублимационной сушкой замороженных растворов в системах с клатратообразованием Огиенко А.Г., Огиенко А.А., Мызь С.А., Юношев А.С., Манаков А.Ю., Болдырева Е.В., Болдырев В.В. Материалы II международной научно-практической конференция "Высокие технологии, фундаментальные и прикладные исследования в физиологии и медицине" |
|
| Использование систем с клатратообразованием для получения высокодисперсного ибупрофена Работа получила третье место на конкурсе работ молодых ученых на конференции «Наукоемкие химические технологии». Е.Г. Зевак, А.Г. Огиенко, Е.В. Болдырева, В.В. Болдырев, А.Ю. Манаков, А.А. Огиенко, С.А. Мызь, А.С. Юношев, Н.В. Кутаев, А.А. Красников IV Молодежная научно-техническая конференция «Наукоемкие химические технологии» |
|
| Генетика природных и экспериментальных популяций дрозофил Захаров И.К., Ваулин О.В., Илинский Ю.Ю., Быков Р.А., Киселёва Е.В., Струнов А.А., Жукова М.В., Коромыслов Ю.А., Васильева Л.А., Антоненко О.В., Волошина М.А., Захаренко Л.П., Иванников А.В., Юрченко Н.Н. Международная конференция «Проблемы популяционной и общей генетики» |
|
| Получение высокодисперсного парацетамола методом вакуумной сушки замороженных растворов в системе ацетон-вода-парацетамол и исследование физико-химических и механических свойств полученного препарата Огиенко А.Г., Болдырева Е.В., Манаков А.Ю., Болдырев В.В., Михайленко М.А., Юношев А.С., Огиенко А.А., Анчаров А.И., Ачкасов А.Ф., Ильдяков А.В., Бурдин А.А., Туманов Н.А., Стопорев А.С., Кутаев Н.В. Семинар с международным участием «Горячие точки химии твердого тела: химия молекулярных кристаллов и разупорядоченных фаз» |
|
| Study of molecular and cytological structure of centromeric regions in Chironomidae Golygina V.V., Broshkov A.D., Ermolaeva O.V., Kiknadze I.I. 18th International Symposium on Chironomidae, Trondheim, Norway, 4-6 July 2011. |
|
| Криптические виды таксона Anopheles messae Fall. (Diptera: Culicidae), их инверсионные и молекулярно-генетические полиморфизмы Ваулин О.В., Новиков Ю.М. Международная конференция “Проблемы популяционной и общей генетики”, посвященная памятной дате – 75-летию со дня рождения академика Ю.П. Алтухова. Москва. 14-16 ноября 2011 г. Международная молодежная конференция “Популяционная генетика: современное состо |
|
| Филогенетический анализ видов рода Diphyllobothrium М.М. Сулейманов, И.И. Брусенцов, Катохин А.В. XLVIII Международная научная студенческая конференция "Студент и научно-технический прогресс” |
|
| Возможность преобладания антропогенного фактора в биогеографии кошачей двуустки Opisthorchis felineus Брусенцов И.И., Катохин А.В., Шеховцов С.В., Мордвинов В.А. Международная конференция «Проблемы популяционной генетики» посвященная памятной дате — 75-летию со дня рождения академиика Ю.П. Алтухова |
|
| 2010 | Investigation of Opisthorchosis felineus transcription profile by direct sequencing cDNA library’s clones S.I. Tatkov, I. I. Brusentsov, M.Yu. Pomaznoy, O.V. Nosareva, A.M. Nayakshin, S. V. Guselnikov, E. V. Brenner, G. V. Vasiliev, A.V. Katokhin, V. A. Mordvinov 7th International conference on bioinformatics of genome regulation and structure\system biology (BGRS/SB'20010). June 20-27, 2010, Novosibirsk |
| Условные мутации у Drosophila melanogaster: образование морфозов Федорова Н.Б., Чадова Е.В., Хоцкина Е.А., Чадов Б.Ф. II Международная конференция "Дрозофила в экспериментальной генетике и биологии", Украина, г. Одесса, 5-11 сентября |
|
| Дарвиновский отбор: способ производства новых видов или перманентное состояние материи? Чадов Б.Ф. IV Международный симпозиум "Эволюция жизни на Земле", 10-12 ноября 2010 г. Томск, Россия |
|
| Использование маркерных ДНК в сравнительной цитогенетике хирономид Гундерина Л.И., Голыгина В.В. V Международная конференция по кариосистематике беспозвоночных животных (KARYO V) с 16 по 20 августа 2010 г. г. Новосибирск |
|
| Молекулярная и цитологическая структура центромерных районов у хирономид Голыгина В.В., Брошков А.Д., Кикнадзе И.И., Карамышева Т.В., Рубцов Н.Б. V Международная конференция по кариосистематике беспозвоночных животных (KARYO V) с 16 по 20 августа 2010 г. г. Новосибирск |
|
| Условные мутации: получение методом морфозов Федорова Н.Б., Чадова Е.В., Хоцкина Е.А., Чадов Б.Ф. VI Международная конференция "Факторы экспериментальной эволюции", Украина,г. Алушта, 21-25 сентября |
|
| Изучение транскриптома Opisthorchis felineus путем секвенирования и функциональной аннотации его к ДНК библиотеке М.Ю.Помазной, И.И. Брусенцов, С.И. Татьков. XLVIII Международная научная студенческая конференция "Студент и научно- технический прогресс", г. Новосибирск, 10-14 апреля 2010 г. |
|
| Диагностика биогельминтозов человека и животных, передающихся через рыбу Брусенцов И.И., Сулейманов М.М., Романов К.В., Катохин А.В., Мордвинов В.А. Конференция "Молекулярная диагностика - 2010" |
|
| Поиск потенциальных мишеней для диагностики паразита Opisthorchis felineus Помазной М. Ю., Татьков С. И., Брусенцов И. И., Сивков А.Ю., Наякшин А.М., Гусельников С.В., Бреннер Е.В., Васильев Г.В., Катохин А.В., Мордвинов В.А. Молекулярная диагностика 22-25 ноября 2010, Москва |
|
| Циклические энергетические потоки и их проявление: материя, живое, сознание Чадов Б.Ф. Первый Международный семинар по БиоКосмологии. 22-25 июля 2010. Великий Новгород |
|
| Генетическая система и мировой энергетический процессинг Чадов Б.Ф. Проблемы экологии. Чтения памяти профессора М.М. Кожова. 20-25 сентября 2010. Иркутск |
|
| Новое прочтение теории естественного отбора Ч.Дарвина Чадов Б.Ф. Проблемы экологии. Чтения памяти профессора М.М. Кожова. 20-25 сентября 2010. Иркутск |
|
| Модель происхождения материи и оценка роли естественного отбора в эволюционном процессе Чадов Б.Ф. Эволюционный синтез и актуальные проблемы эволюции человека ( к 110-летию Ф.Г. Добржанского) 11-12 июня 2010. С.Петербург. |
|
| An experimental and computation approach to search for the transcription factor GAGA binding sites Omelina E.S., Oshchepkov D.Yu., Baricheva E.M., Merkulova T.I. BGRS-2010 |
|
| Диагностика биогельминтозов человека и животных, передающихся через рыбу Брусенцов И.И., Сулейманов М.М., Романов К.В., Катохин А.В., Мордвинов В.А. VII Всероссийская научно-практическая конференция с международным участием «Молекулярная диагностика - 2010» |
|
| Перспективы использования молекулярно-генетических методов исследований в диагностике описторхидозных инвазий А.В. Катохин, А.В. Евтушенко, И.И. Брусенцов, А.С. Солодянкин, Б.В. Ромашов, В.А. Мордвинов Мiжнародная науково-практичная конференция «Сучаснi системи биобезпеки та биозахисту у ветеринарнiй медицинi» |
|
| 2009 | Conditional dominant lethals - a new class of mutations in Drosophila Chadov B.F. 3-rd Annual Wold Congress of Gene-2009. December 1-7, 2009.Foshan (China) |
| Анализ влияния мутаций по гену Trithorax – like на формирование глаза Drosophila melanogaster Н.В. Павлова, Д.А. Карагодин 5-й съезд ВОГИС, посвященный 200-летию со дня рождения Чарльза Дарвина, Москва, 21-28 июня 2009 г. |
|
| Молекулярно-генетическая филогения видов семейства Opisthorchiidae А.В. Катохин, С.В. Шеховцов, К.В. Романов, И.И. Брусенцов, Н.И. Юрлова, В.А. Мордвинов III Межрегиональная конференция паразитологов Сибири и Дальнего Востока, посвященная 80-летию профессора К.П.Федорова // Новосибирск. 15-20 сентября 2009 года |
|
| Opisthorchis felineus and Metorchis bilis: development of ITS-2 based multiplex PCR assay for their identification and discrimination I. I. Brusentsov, A.V. Katokhin, M.N. Lvova, Z. Sacharovskaja, A. E. Sazonov, L.M. Ogorodova, O. Fedorova, N.A. Kolchanov, V.A. Mordvinov Joint International Tropical Medicine Meeting 2009 (6th Seminar on Food- and Water-borne Parasitic Zoonoses (FBPZ6)) |
|
| Генетические мутации, подготавливающие процесс видообразования Федорова Н.Б., Чадова Е.В., Хоцкина Е.А.,Чадов Б.Ф. V Международная конференция " Факторы экспериментальной эволюции организмов" 21-25 сентября 2009. Алушта. Украина. |
|
| Циклическое движение как способ генерации материального Чадов Б.Ф. V Российский философский конгресс.25-28 августа 2009. Новосибирск |
|
| Interspecific hybridization between sibling species of Chironomus plumosus group ((Diptera: Chironomidae) Golygina V.V., Istomina A.G., Kiknadze I.I. XVII International Symposium on Chironomidae |
|
| Энергетическое предназначение живого и генетическая система Чадов Б.Ф. Международная конференция "Хромосома 2009", август 31- сентябрь 6, 2009. Новосибирск |
|
| Изменение энергетического статуса дрозофилы в результате генетической мутации Федорова Н.Б., Д.В. Петровский, Е.В. Чадова, Е.А. Хоцкина, М.П. Мошкин, Б.Ф. Чадов Пятый Съезд Вавиловского общества генетиков и селекционеров Москва, 21-28 июня 2009 |
|
| Эволюция как энергетический процесс: генетическое изучение энергетической стороны эволюции Чадов Б.Ф. Пятый Съезд Вавиловского общества генетиков и селекционеров Москва, 21-28 июня 2009 |
|
| Концепция естественного отбора Ч.Дарвина и её состояние на начало XXI века Чадов Б.Ф. Синтетическая теория эволюции: состояние, проблемы, перспективы (Международная научная конференция, посвященная 200-летию со дня рождения Ч.Дарвина). Украина. Луганск. 15-19 июня 2009 |
|
| Энергетическое предназначение живого и эволюционный процесс Чадов Б.Ф., Федорова Н.Б. Чарльз Дарвин и современная наука, 21-25 сентября, Ст.Петербург |
|
| Полиморфизм мультилокусных маркёров ДНК и генных последовательностей в популяциях Drosophila melanogaster Ваулин О.В., Захаров И.К. Съезд генетиков и селекционеров, посвященный 200-летию со дня рождения Ч. Дарвина. 5 съезд Вавиловского общества генетиков и селекционеров. Москва, 31-28 июня 2009 г. Часть 2. – Москва. 2009. С. 153. |
|
| Исследование строения Wolbachia и их влияние на продолжительность жизни линий Drosophila melanogaster, несущих мутации по гену Trithorax-like Бочериков А.М., Воронин Д.А., Баричева Э.М., Захаров И.К., Киселева Е.В. Съезд генетиков и селекционеров, посвященный 200-летию со дня рождения Ч. Дарвина. 5 съезд Вавиловского общества генетиков и селекционеров. Москва, 21-28 июня 2009 г. Часть 2. – Москва. 2009. С. 204. |
|
| Динамические процессы и генетическая изменчивость в природных популяциях Захаров И.К., Ваулин О.В., Илинский Ю.Ю., Синянский Я.Я., Коромыслов Ю.А. , Коваленко Л.В. , Иванников А.В., Захаренко Л.П., Волошина М.А., Чересиз С.В. , Юрченко Н.Н. Съезд генетиков и селекционеров, посвященный 200-летию со дня рождения Ч. Дарвина. 5 съезд Вавиловского общества генетиков и селекционеров. Москва, 21-28 июня 2009 г. Часть 2. – Москва. 2009. С. 161. |
|
| Динамические процессы в генофондах природных популяций дрозофил. Захаров И.К., Илинский Ю.Ю., Ваулин О.В., Синянский Я.Я., Иванников А.В., Волошина М.А., Захаренко Л.П., Юрченко Н.Н. В кн.: Чарльз Дарвин и современная наука. Сборник тезисов международных научных конференций “Чарльз Дарвин и современная биология” (21-23 сентября 2009 г., Санкт-Петербург) и “Теория эволюции: между наукой и идеологией. Историко-научные и философско-метод |
|
| Анализ влияния мутаций по гену Trithorax – like на формирование глаза Drosophila melanogaster Н.В. Павлова, Д.А. Карагодин 5-й съезд ВОГИС, посвященный 200-летию со дня рождения Чарльза Дарвина, Москва, 21-28 июня 2009 г. 2009, Ч. 2. С. 381 |
|
| Изменение энергетического статуса дрозофилы в результате генетической мутации Федорова Н.Б., Петровский Д.В.,Чадова Е.В., Хоцкина Е.А., Мошкин М.П.,Чадов Б.Ф. V Съезд Вавиловского общества генетиков и селекционеров |
|
| Исследование генетического разнообразия возбудителей биогельминтозов человека, передающихся через рыбу, ракообразных и продукты их переработки В.А. Мордвинов, А.В. Катохин, И.И. Брусенцов, К.В. Романов, С.В. Шеховцов, С.И. Татьков, Д.П.Фурман, А.Ю.Сивков, М.Ю.Помазной, М.Н.Львова, М.Ю.Шаманина, К.С.Задесенец, Н.Б.Рубцов. Итоговая конференция по приоритетному направлению "Живые системы" за 2009 год |
|
| 2008 | Исследование влияния мутаций гена Trithorax-like на расхождение Х-хромосом в мейозе самок Drosophila melanogaster Лаухина О.В., Федорова С.А., Баричева Э.М. I Международная конференция "Дрозофила в экспериментальной генетике и биологии" |
| Исследование регуляторного потенциала второго интрона гена Trithorax-like Drosophila melanogaster Федорова Е.В., Карагодин Д.А., Баричева Э.М. I Международная конференция "Дрозофила в экспериментальной генетике и биологии", 15-20 сентября 2008 г., Харьков, Украина |
|
| Взаимодействие между SUUR и HP1 обуславливает позднюю репликацию интеркалярного гетерохроматина у Drosophila Пиндюрин А. В., Шлома В.В., Болдырева Л.В., Похолкова Г.В., Колесникова Т.Д., Белякин С.Н., Беляева Е.С., Жимулев И.Ф. IV съезд Российского Общества биохимиков и молекулярных биологов, 10-15 мая, Новосибирск, 2008 |
|
| Interaction between SUUR and HP1 determines late replication timing of intercalary heterochromatin in Drosophila Pindyurin A.V., Shloma V.V., Boldyreva L.V., Pokholkova G.V., Kolesnikova T.D., Kozhevnikova E.N., Belyakin S.N., Belyaeva E.S., Zhimulev I. F. The Sixteenth International Workshop on the MOLECULAR AND DEVELOPMENTAL BIOLOGY OF DROSOPHILA (CRETE XVI EMBO Conference). Kolymbari, Crete (Greece), June 22-28, 2008 |
|
| Цитогенетические исследования хирономид Горного Алтая Кикнадзе И.И., Истомина А.Г., Голыгина В.В., Брошков А.Д. Биоразнообразие, проблемы экологии Горного Алтая и сопредельных регионов: настоящее, прошлое, будущее. 22-26 сентября 2008 г. Горно-Алтайск |
|
| Механизмы генетической изменчивости популяций и видов Diptera: Полиморфизм и инсерционные мутации в различных условиях Васильева Л.А., Ваулин О.В., Илинский Ю.Ю., Захаренко Л.П., Антоненко О.В., Перепёлкина М.П., Коромыслов Ю.А., Волошина М.А., Иванников А.В., Чересиз С.В., Юрченко Н.Н., Захаров И.К. Программа фундаментальных исследований РАН. № 11. “Биоразнообразие и динамика генофондов”. Подпрограмма 2 “Динамика генофондов”. Материалы отчётной конференции. – Москва. 2008. С. 24-25 |
|
| Мутация в онтогене дрозофилы вызывает перемещение ретротранспозона 412 Федорова Н.Б., Чадова Е.В. IV международная конференция "Факторы экспериментальной эволюции организмов" |
|
| Условные доминантные летали у дрозофилы: высокая двигательная активность имаго Федорова Н.Б., Чадов Б.Ф. Вторые чтения, посвященные памяти В.И. Корогодина и В.А. Шевченко «Актуальные вопросы генетики, радиобиологии и радиоэкологии» |
|
| 2007 | High resolution analysis of heterochromatin organization using SuUR Su(var)3-9 double mutants in Drosophila melanogaster E.N. Andreyeva, T.D. Kolesnikova, O.V. Demakova, S.A. Demakov, M. Mendez-Lago, G.V. Pokholkova, E.S. Belyaeva, U.A. Komor, A.V. Doroshkov, F. Rossi, P. Dimitri, A. Villasante, I.F. Zhimulev 8th International Conference on Drosophila Heterochromatin, Gubbio, 3 – 9 June 2007 |
| Организация эпигенетически репрессированных районов хромосом Drosophila melanogaster Колесникова Т.Д., Андреева Е.Н., Пиндюрин А.В., Ананько Н.Г., Белякин С.Н., Шлома В.В., Юрлова А.А, Макунин И.В., Похолкова Г.В., Волкова Е.И., Заруцкая Е.А., Кокоза Е.Б., Семешин В.Ф., Беляева Е.С., Жимулёв И.Ф. Current Evolutionary Thinking in Biology, Medicine and Sociology. International Conference Dedicated to 90 Anniversary of Prof. Dmitry K. Belyaev 7-9 August 2007, Novosibirsk, Russia |
|
| Белок SUUR и его действие на структуру политенных хромосом И.Ф.Жимулёв, Е.С.Беляева, Е.Н.Андреева, О.В.Демакова, Г.В.Похолкова, А.В. Пиндюрин II Cъезд Общества клеточной биологии совместно с Юбилейной конференцией, посвященной 50-летию Института цитологии РАН. Санкт-Петербург, 16 - 19 октября 2007 |
|
| The SUUR protein affects late replicating chromatin domains in Drosophila melanogaster Andreyeva E., Pindyurin A., Kolesnikova T., Shloma V., Demakova O., Boldyreva L., Pokholkova G., Belyaeva E., Zhimulev I Meeting on EUKARYOTIC DNA REPLICATION & GENOME MAINTENANCE, September 5-9, 2007. Cold Spring Harbor Laboratory, Cold Spring Harbor, New York |
|
| Геномное картирование генов-мишеней белка SUUR Drosophila melanogaster Пиндюрин А.В. Международная молодежная научно-методическая конференция "Проблемы молекулярной и клеточной биологии", г. Томск |
|
| Механизмы генетической изменчивости популяций и видов Diptera: Разнообразие геномов Захаров И.К., Кикнадзе И.И., Киселева Е.В., Гундерина Л.И., Ваулин О.В., Синянский Я.Я., Илинский Ю.Ю., Воронин Д.А., Жукова М.В., Бочериков А.М., Голыгина В.В., Истомина А.Г., Вайсман Н.Я., Иванников А.В. Программа фундаментальных исследований РАН. № 11. “Биоразнообразие и динамика генофондов”. Подпрограмма 2. “Динамика генофондов”. Отчетная конференция. |
|
| Механизмы генетической изменчивости популяций и видов Diptera: Взаимодействия генетических и средовых факторов Захаров И.К., Киселёва Е.В., Илинский Ю.Ю., Воронин Д.А., Ваулин О.В., Жукова М.В., Бочериков А.М., Вайсман Н.Я., Синянский Я.Я., Иванников А.В. Конференция “Биоразнообразие и динамика генофондов”, посвященная памяти академика Ю.П. Алтухова |
|
| Изменчивость фрагментов геномной ДНК в североевразийских популяциях Drosophila melanogaster Ваулин О.В., Захаров И.К. Материалы конференции. “Современные проблемы биологической эволюции”. К 100-летию Государственного Дарвиновского музея. 17-20 сентября 2007. Москва. - М.: Изд-во ГДМ. 2007. С. 167-168. |
|
| Механизмы генетической изменчивости популяций и видов Diptera: Транспозиции мобильных генетических элементов при различных воздействиях Васильева Л.А., Захаренко Л.П., Коваленко Л.В., Антоненко О.В., Фёдорова Н.Б., Перепёлкина М.П., Волошина М.А., Коромыслов Ю.А., Юрченко Н.Н., Чересиз С.В., Захаров И.К. Программа фундаментальных исследований РАН. № 11. “Биоразнообразие и динамика генофондов”. Подпрограмма 2 “Динамика генофондов”. Материалы отчётной конференции, посвящённой памяти академика Ю.П. Алтухова. – Москва. 2007. С. 41. |
|
| Анализ влияния нового гипоморфного аллеля гена Trithorax – like (Trl en82) на структуру глаза и оогенез Drosophila melanogaster Н.В. Павлова, Д.А. Карагодин Проблемы молекулярной и клеточной биологии. Международная молодежная научно-методическая конференция. 9-12 мая 2007г. |
|
| Новый взгляд на роль хромосомной перестройки в видообразовании Федорова Нина Борисовна Современные проблемы биологической эволюции. К 100-летию Государственного Дарвиновского музея |
|
| Онтогенез, онтогены и синдром геномной несовместимости. Клеточные, молекулярные и эволюционные аспекты морфогенеза Чадов Б.Ф., Федорова Н.Б.,Хоцкина Е.А., Чадова Е.В. Симпозиум с международным участием «Клеточные, молекулярные и эволюционные аспекты морфогенеза» 9-11 октября 2007. Москва |
|
| Геномная нестабильность в линиях дрозофилы с факультативными доминантными леталями Федорова Н.Б. Международная молодежная научно-методической конференция "Проблемы молекулярной и клеточной биологии" |
|
| Межвидовая гибридизация видов-близнецов группы Chironomus plumosus (Diptera, Chironomidae) Голыгина В.В., Истомина А.Г., Кикнадзе И.И. XIII С Ъ Е З Д РУССКОГО ЭНТОМОЛОГИЧЕСКОГО ОБЩЕСТВА Краснодар, 9 - 15 сентября 2007 г. |
|
| 2006 | Квазицикл " Ген-проген" -имманентное свойство живой системы Чадов Б.Ф. IV Международная конференция " Проблема вида и видообразование". 25-28 сентября 2006. г. Томск. Томский Государственный Университет |
| Особенности структуры кариотипа в роде Propsilocerus Kieff. (Diptera, Chironomidar) Кикнадзе И.И., Истомина А.Г., Шин Ву, Голыгина В.В. IV Международная конференция по кариосистематике беспозвоночных животных |
|
| Цитогенетическая дивергенция геномов видов-близнецов группы Chironomus plumosus (Diptera:Chironomidae) Голыгина В.В., Кикнадзе И.И., Истомина А.Г., Гундерина Л.И., Мирошниченко Л.А., Гусев В.Д. IV Международная конференция по кариосистематике беспозвоночных животных |
|
| Мутация в онтогене и радиационно-индуцированная нестабильность у Drosophila melanogaster Федорова Н.Б. V Съезд по радиационным исследованиям (радиобиология, радиоэкология, радиационная безопасность. 10-14 апреля 2006, Москва. |
|
| Эволюция геномной ДНК у видов рода Chironomus (Diptera, Chironomidae) Гундерина Л.И., Кикнадзе И.И., Истомина А.Г., Голыгина В.В., Гадыльшина Е.Г. VII Межрегиональное совещание энтомологов Сибири и Дальнего Востока в рамках Сибирской зоологической конференции |
|
| Дивергенция последовательностей дисков и филогения видов-близнецов в группе plumosus (Diptera, Chironomidae) Голыгина В.В., Кикнадзе И.И., Гундерина Л.И., Истомина А.Г., Егорова Е.М., Мирошниченко Л.А., Гусев В.Д. VII Межрегиональное совещание энтомологов Сибири и Дальнего Востока в рамках Сибирской зоологической конференции Новосибирск, 20-24 сентября 2006 г. |
|
| Мутации в онтогенах Drosophila melanogaster и проблема стерильности Чадов Борис Генетика репродукции насекомых и её практическое применение, 24-25 апреля, г. Москва (швейцарско- российская международная конференция) |
|
| Мутации в онтогене переводят геном из стабильного состояния в нестабильное Федорова Н.Б., Васильева Л.А.,Чадов Б.Ф. Международная конференция "Генетика в России и мире", посвященная 40-летию Института общей генетики им. Н.И. Вавилова РАН. 28 июня-2 июля 2006. Москва |
|
| Дивергенция геномной ДНК у видов-двойников группы Chironomus plumosus (Diptera:Chironomidae) Гундерина Л.И., Кикнадзе И.И., Истомина А.Г., Голыгина В.В., Гадыльшина Е.Г. IV Международная конференция по кариосистематике беспозвоночных животных, 28-30 августа 2006 г. Санкт-Петербург |
|
| Анализ механизмов влияния гена Trithorax-like на нарушение процесса быстрого транспорта в оогенезе Drosophila melanogaster А.А. Огиенко, Д.А. Карагодин, С.А. Федорова, Э.М. Баричева Международная конференция “Генетика в России и мире” |
|
| Механизмы генетической изменчивости популяций и видов Diptera: Мобильные генетические элементы в популяциях и в опыте Васильева Л.А., Захаренко Л.П., Коваленко Л.В., Чмуж Е.В., Шестакова Л.А., Волкова В.С., Выхристюк О.В., Антоненко О.В., Фёдорова Н.Б., Перепёлкина М.П., Волошина М.А., Коромыслов Ю.А., Юрченко Н.Н., Захаров И.К. Программа фундаментальных исследований РАН. № 11. “Биоразнообразие и динамика генофондов”. Подпрограмма 2. “Динамика генофондов”. Сборник материалов. – М: ФИАН. 2006. С. 24-25. |
|
| Механизмы генетической изменчивости популяций и видов Diptera: Разнообразие геномов Захаров И.К., Кикнадзе И.И., Киселёва Е.В., Гундерина Л.И., Ваулин О.В., Синянский Я.Я., Илинский Ю.Ю., Воронин Д.А., Жукова М.В., Бочериков А.М., Голыгина В.В., Истомина А.Г., Вайсман Н.Я., Иванников А.В. Программа фундаментальных исследований РАН. № 11. “Биоразнообразие и динамика генофондов”. Подпрограмма 2. “Динамика генофондов”. Сборник материалов. – М: ФИАН. 2006. С. 26-28. |
|
| Структура гибридной зоны между кедром сибирским и кедровым стлаником в Северном Прибайкалье: полевые наблюдения и молекулярно-генетический анализ Попов А.Г., Ваулин О.В., Ван Джун. Фундаментальные проблемы новых технологий в третьем тысячелетии. Материалы 3-й Всероссийской конференции молодых ученых. Томск. 3-6 марта 2006 г. – Томск: Изд-во Института оптики атмосферы СО РАН. 2006. С. 544-548. |
|
| Разнообразие мультилокусных маркёров генома Drosophila melanogaster в популяциях Палеарктики Ваулин О.В. Биология - наука 21 века. 10-я Пущинская школа-конференция молодых учёных, посвященная 50-летию Пущинского НЦ РАН. Пущино, 17-21 апреля 2006 г. Сборник тезисов. – Пущино: Пущинский научный центр РАН. 2006. С. 263. |
|
| Временнáя динамика структуры генофонда в популяции Drosophila melanogaster Умань (Украина) Ваулин О.В., Жариков Т.Ю. Биология - наука 21 века. 10-я Пущинская школа-конференция молодых учёных, посвященная 50-летию Пущинского НЦ РАН. Пущино, 17-21 апреля 2006 г. Сборник тезисов. – Пущино: Пущинский научный центр РАН. 2006. С. 264. |
|
| Молекулярно-генетическая характеристика нового гипоморфного аллеля гена Trithorax – like Drosophila melanogaster и анализ его взаимодействия с геном lozenge Н.В. Павлова XLIV международная научная студенческая конференция «Студент и научно-технический прогресс» |
|
| Анализ механизмов влияния гена Trithorax-like на нарушение процесса быстрого транспорта в оогенезе Drosophila melanogaster Огиенко А.А., Карагодин Д.А., Федорова С.А., Лашина В.В., Федорова Е.В., Баричева Э.М. Материалы 10-ой Пущинской Школы-конференции молодых ученых |
|
| 2001 | Космос, космология и биокосмология (нео-Аристотелизм Чадов Борис Федорович Второй Международный семинар по БиоКосмологии |
Гранты
| 2020 | Механизмы регуляции программируемой гибели клеток эукариот посредством аутофагии РФФИ, номер гранта 20-04-00496 А |
| 2019 | Популяционная геномика и эволюционная история комаров-переносчиков заболеваний человека в России РНФ, номер гранта 19-14-00130 |
| 2014 | Сетчатая эволюция рода Aquilegia (Ranunculaceae) и ее таксономические последствия РФФИ, номер гранта 14-04-01415 |
| Изучение эволюции геномов видов рода Chironomus c использованием молекулярных маркеров на основе ядерной и митохондриальной ДНК РФФИ, номер гранта 14-04-01126 |
|
| 2012 | «Дизайн комбинированных нанокомпозитов, не требующих использования фреонов в составе противоастматических лекарственных форм» Правительство новосибирской области на проведение прикладных научных исследований |
| Исследование генетических систем, контролирующих динамику и реорганизацию актинового цитоскелета в клетках эукариот, на примере оогенеза Drosophila melanogaster РФФИ, номер гранта 12-04-31804 мол_а |
|
| Филогенетические исследования комаров рр. Aedes и Ochlerotatus (Diptera: Culicidae) Научный проект Храбровой Натальи Валерьевны из НИИ биологии и биофизики Томского государственного университета, г. Томск, в Институте цитологии и генетики СО РАН, г. Новосибирск РФФИ, номер гранта 12-04-90826-мол_рф_нр |
|
| Идентификация генов-мишеней транскрипционного фактора GAGA, контролирующих формирование дорзальных выростов хориона на яйцах Drosophila melanogaster ООО "ОПТЭК" |
|
| «Дизайн фармацевтических субстанций с использованием крио-нанотехнологий» ОПТЭК |
|
| Интеграционный проект СО РАН №108 "Нетрадиционные способы введения лекарственных веществ в организм и физические методы модифицирования их свойств" Номер гранта №108 |
|
| Динамичность генофондов, генетическая и фенотипическая изменчивость популяций РФФИ, номер гранта 12-04-01319-а |
|
| 2011 | Научная работа Артемова Глеба Николаевича из Научно-исследовательского института биологии и биофизики Томского государственного университета, г. Томск, в Институте цитологии и генетики СО РАН, г. Новосибирск. Разработка детальной цитогенетической и физической карты малярийного комара Anopheles atroparvus Thiel РФФИ, номер гранта 11-04-90801-моб_ст |
| Грант Президиума РАН (2009-2011): Программа фундаментальных исследований РАН “Биоразнообразие и динамика генофондов”, подпрограмма “Динамика генофондов ” (руководитель Захаров И.К.); Президиум РАН, номер гранта №1-13/10 |
|
| 2010 | Идентификация генов-мишеней транскрипционных факторов, участвующих в контроле развития глаза дрозофилы РФФИ, номер гранта 10-04-01011 |
| Идентификация видов и изучение эволюции геномов с помощью молекулярных маркеров (на примере видов рода Chironomus) РФФИ, номер гранта 10-04-00899 |
|
| 2009 | Участие в конференции BIT Life SciencesУчастие в конференции BIT Life Sciences' 3rd Annual World Congress of Gene-(2009)(WCG-2009) РФФИ, номер гранта 09-04-08258-з |
| Участие в V Международной научной конференции «Факторы экспериментальной эволюции организмов» РФФИ, номер гранта 09-04-08300-з |
|
| Изучение молекулярной организации центромерной ДНК и её роли в процессе видообразования у хирономид Номер гранта 09-04-01440a |
|
| Генетические факторы, влияющие на изменчивость генофондов популяций дрозофил РФФИ, номер гранта 09-04-00872а |
|
| 2008 | Свойства генов, управляющих онтогенезом дрозофилы РФФИ, номер гранта 08-04-00094-а |
| Генофонды и генетическое разнообразие природных и экспериментальных популяций Президиум РАН, номер гранта 26.30 |
|
| 2006 | Исследование изменчивости рисунка мобильных генетических элементов (МГЭ) дрозофилы при различных стрессовых воздействиях РФФИ, номер гранта 06-04-48116-а |
| Изучение регуляции экспрессии гена Trithorax-like и его роли в онтогенезе D. melanogaster РФФИ, номер гранта 06-04-49000 |
|
| Грант Президиума РАН (2006-2007): Программа фундаментальных исследований РАН “ динамика генофондов”, “Динамика генофондов растений, животных и человека” №11.4.1 (руководитель Захаров И.К.); Президиум РАН, номер гранта №11.4.1 |
|
| 2005 | Инсерционный мутагенез, как фактор микроэволюции в природных популяциях дрозофил РФФИ, номер гранта 05-04-48838а |
| 2004 | Гены, управляющие развитием дрозофилы: новые мутации РФФИ, номер гранта 04-04-48100-а |
| Изучение механизма возникновения сайт-направленных мутаций в протоонкогенах семейства ras РФФИ, номер гранта 04-04-48136 |
Научное руководство
| 2023 | Регуляция запрограммированной гибели клеток в слюнных железах Drosophila melanogaster на фоне недостатка транскрипционного фактора GAGA Сюй Чжихуа 2023-06-10 |
| 2017 | Эффекты введения дексаметазона в раннем возрасте на паттерн метилирования ДНК в гиппокампе мышей Садчиков Артем александрович , 2017-06-14 |
| 2012 | Изменчивость по фрагменту гена Adar у видов малярийных комаров А и В Anopheles messeae и Anopheles beklemishevi Алейникова Софья Александровна , 2012-09-25 |
| 2010 | ПОЛУЧЕНИЕ И АНАЛИЗ МУТАЦИЙ, ЗАТРАГИВАЮЩИХ ВТОРОЙ ИНТРОН ГЕНА TRITHORAX-LIKE DROSOPHILA MELANOGASTER Федорова Елена Владимировна , 2010-11-30 |
| Полиморфизм некодирующих последовательностей ДНК хлоропластов Aegilops tauschii Хусаинова Рината Равильевна 2010-06-02 |
|
| 2009 | Идентификация и анализ сайтов связывания транскрипционного фактора GAGA, кодируемого геном Trithorax-like Drosophila melanogaster. Омелина Евгения Сергеевна , 2009-06-04 |
| 2008 | МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКАЯ ХАРАКТЕРИСТИКА МУТАЦИЙ ГЕНА TRITHORAX-LIKE И ИХ ВЛИЯНИЕ НА ООГЕНЕЗ DROSOPHILA MELANOGASTER Огиенко Анна Александровна , 2008-10-01 |
| 2007 | ДЕЛЕЦИОННОЕ КАРТИРОВАНИЕ 5’-РЕГУЛЯТОРНОЙ ОБЛАСТИ ГЕНА Trithorax – like Drosophila melanogaster Сокуева Наталья Александровна , 2007-06-01 |
| 2006 | МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКАЯ ХАРАКТЕРИСТИКА НОВОГО ГИПОМОРФНОГО АЛЛЕЛЯ ГЕНА Trithorax – like И АНАЛИЗ ЕГО ВЛИЯНИЯ НА СТРУКТУРУ ГЛАЗА И ООГЕНЕЗ Drosophila melanogaster Батулина Надежда Викторовна , 2006-06-05 |
Патенты
| 2013 | Способ получения высокодисперсного ибупрофена Болдырева Е.В., Огиенко А.Г., Болдырев В.В., Манаков А.Ю., Мызь С.А., Огиенко А.А., Зевак Е.Г., Ильдяков А.В., Шахтшнейдер Т.П. Номер патента 2491919 |
| 2011 | Способ получения высокодисперсного парацетамола Огиенко А.Г., Болдырева Е.В., Манаков А.Ю., Болдырев В.В., Михайленко М.А., Юношев А.С., Огиенко А.А., Анчаров А.И., Ачкасов А.Ф., Ильдяков А.В., Бурдин А.А., Туманов Н.А., Стопорев А.С., Кутаев Н.В. Номер патента заявка №2010152150 от 20.12.2010 |
| © 2010-2026 ИЦиГ СО РАН. Все права защищены. |