

Система учета научной деятельности (ASSA) |


|
Лаборатория популяционной этногенетики (т.56)Отдел молекулярной генетики человека (т.140)Научные результаты Сотрудники О Подразделении 1. Основное направление фундаментальных и фундаментально-ориентированных исследований Комплексное изучение разнообразия генофондов коренных этносов Сибири в эволюционном, популяционном и биомедицинском аспектах. Задачи, решаемые в рамках основного направления на данном этапе:
2. Аннотация базового бюджетного проекта подразделения Проект: VI.58.1.1. Особенности формирования молекулярно-генетических механизмов метаболических нарушений и наследственных заболеваний у жителей Северной Азии и разработка новых подходов к их коррекции (№ гос. Регистрации 01201280318). Руководитель – член-корреспондент РАМН, д.м.н. Воевода М.И. Лаборатория популяционной этногенетики (т.56) отвечает за Блок №3 бюджетного проекта «Оценка состояния генофонда бурят Усть-Ордынского Бурятского округа Иркутской области».
3. Прикладные разработки Прикладных разработок нет.
4. Иллюстрированное описание лучших результатов, полученных подразделением за последние 5 лет Впервые было показано, что у народов Западной Берингии (северо-востока России) и во всех исследованных популяциях коренных жителей Америки присутствует уникальный аллель микросателлитного локуса D9S1120 (с 9-ю повторами), который не был найден в других мировых популяциях (Рис. 1). Это позволило предположить, что все коренные жители Америки произошли от одной предковой сибирской популяции. Такое заключение подразумевает, что все обнаруженные копии 9-повторного аллеля имеют общее происхождение и что на распространение этого аллеля не влиял естественный отбор. Чтобы проверить эти предположения, были исследованы 34 однонуклеотидных полиморфных сайта (SNP) на расстоянии порядка 500 т.п.н. вокруг D9S1120 в 21 популяциях коренных жителей Америки и Западной Берингии, а также в 54 других популяциях со всего мира. Все хромосомы с 9-повторным аллелем имеют очень похожие гаплотипы в окрестностях D9S1120, из чего следует, что все они происходят из одного источника. Девяносто один процент таких хромосом несут один и тот же гаплотип длиной 76.26 т.п.н., который был назван нами «Американский Гаплотип» (АГ). Три наблюдения позволяют заключить, что повсеместное распространение и высокая частота 9-повторного аллеля, наблюдаемые в популяциях Америки, не являются результатом положительного отбора. Во-первых, сам по себе АГ не встречается в Америке с высокой частотой. Во-вторых, АГ не отличается по частоте от других гаплотипов такой длины. В-третьих, в метисных популяциях Латинской Америки локус D9S1120 не отличается от других микросателлитных участков по доле аллелей, унаследованных от индейских предков. С помощью нового метода определения времени до последнего общего предка всех копий аллеля, основанного на оценке числа прошедших поколений, было вычислено время до последнего общего предка для 9-повторного аллеля, которое составило от 7 325 до 39 900 лет, в зависимости от используемой демографической модели. Полученные результаты подтверждают гипотезу, что все коренные жители Америки и Западной Берингии унаследовали большую часть генов от единственной предковой популяции, которая была изолирована от других популяций Азии перед переселением на Американский континент. (Mol Biol Evol 2009. 26(5):995-1016)
Рисунок 1. Распределение частоты аллеля 9RA D9S1120 в популяциях Сибири, Азии и Америки
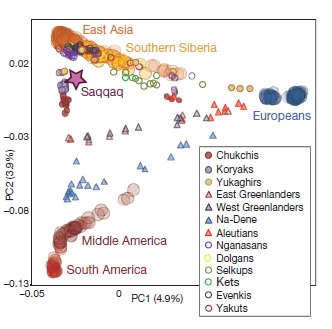
В данной работе впервые изучен геном мужчины, жившего около 4 000 лет назад в Гренландии и принадлежавшего к самой древней из найденных там археологических культур (Саккак). ДНК была выделена из образцов волос, сохранившихся в вечной мерзлоте. Было определено 79% последовательности генома со средним покрытием 20х, что близко к пределу возможностей современных технологий секвенирования. С высокой точностью были определены 353 151 однонуклеотидных полиморфных сайта (SNP), 6.8% из которых до этого не были опубликованы. Оцененная контаминация ДНК составила не более 0.8%. Анализ функционально значимых замен позволил составить фенотипический портрет человека, о котором получено мало информации из археологического материала (человек Саккак имел тёмный цвет волос и смуглый цвет кожи, группу крови А (II), положительный резус-фактор и был адаптирован к холодному климату). Сравнение SNP генома древнего человека и представителей современных этносов выявило популяции, которые наиболее близки к исследуемому образцу – ими оказались чукчи и коряки (Рис. 2). Полученные данные подтверждают гипотезу о дополнительной волне миграции из Сибири в Новый Свет около 5 500 лет назад, независимо от тех волн, которые дали начало популяциям индейцев и инуитов (Опубликовано: Nature 2010. 463(7282):757-62).
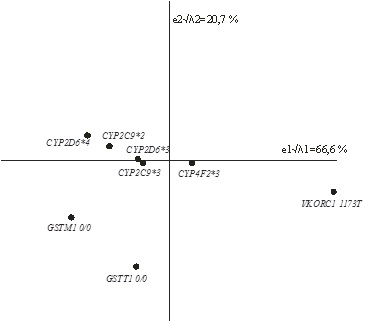
У представителей самодийских этносов (селькупов, лесных ненцев, нганасан) впервые проведено исследование полиморфизма генов системы биотрансформации ксенобиотиков CYP2D6 (аллели CYP2D6*3, CYP2D6*4), с («нулевые» генотипы GSTМ1 0/0 и GSTT1 0/0), рассматриваемых в качестве генетических маркёров риска онкологических заболеваний. Исследование выявило существенную вариабельность частот распределения CYP2D6*4 и GSTМ1 0/0 в северных популяциях. В то же время, частоты остальных вариантов (CYP2D6*3 и GSTT1 0/0) достоверно не отличаются между собой в коренных популяциях селькупов, лесных ненцев и нганасан. По частотам аллеля CYP2D6*4 коренные этносы занимают промежуточное положение между русскими Сибири и монголоидами Китая. Однако суммарная представленность нулевых генотипов GSTМ1 0/0 и GSTT1 0/0 у коренных этносов достоверно ниже, чем в популяциях русских Сибири и монголоидов Китая (р<0.05). На основании частот полиморфных вариантов этих генов выдвинуто предположение о пониженном риске онкологических заболеваний у коренных самодийских этносов по сравнению с русскими Сибири. Исключение составляет популяция лесных ненцев, у которых зарегистрирована повышенная частота генотипов GSTМ1 0/0, что может быть обусловлено своеобразием брачной структуры и повышенным коэффициентом инбридинга, характерным для лесных ненцев. Полученные результаты могут также иметь важное значение при прогнозировании вероятности осложнений при использовании лекарственных препаратов, которые метаболизируются ферментами GSTM1, GSTT1 и CYP2D6. (Опубликовано: Вавиловский журнал генетики и селекции 2011. Т. 15. № 3. С. 585-598).
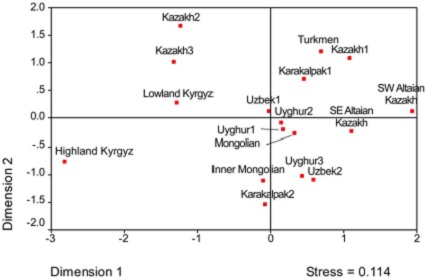
Для того чтобы прояснить популяционную историю казахского этноса по отцовской линии, впервые был проведён анализ нерекомбинирующего участка Y-хромосомы в популяциях казахов, проживающих в южном Алтае, по 60 биаллельным маркёрам и 17 STR (коротким тандемным повторам). Были обнаружены сильные различия в материнской и отцовской наследственности казахов. Притом что в их популяциях обнаруживается большое разнообразие восточно- и западно-евразийских гаплогрупп митохондриальной ДНК (мтДНК), только три Y-гаплогруппы присутствуют со значимой частотой (C3*, C3c и O3a3c*). По данным анализа Y-STR, внутри этих трёх гаплогрупп число генетических вариантов не велико. Близкое родство всех казахов по отцовской линии выделяет их среди других народов Центральной Азии. Комбинированный анализ данных по биаллельным маркёрам и Y-STR выявил различия между популяциями казахов из разных областей Центральной Азии (Рис 4). Различия между казахами Казахстана и Алтая не были вызваны смешением последних с другими народами Алтая. Также исследованные популяции казахов несут следы генетического смешения времён Монгольской Империи 13-го века. В целом можно сказать, что генетический профиль современных казахов обусловлен их историческими социально-культурными традициями и действием факторов микроэволюции. (Опубликовано: PLoS One 2011. 6(3):e17548).
Впервые на представительной выборке был проанализирован полиморфизм митохондриальной ДНК (мтДНК) и Y-хромосомы у представителей северных и южных алтайских популяций с целью выявления генетических взаимосвязей между народами Алтая и другими этносами. МтДНК исследовалась с помощью анализа ПДРФ и секвенирования контрольной области. Нерекомбинирующие участки Y-хромосомы были прогенотипированы с использованием более 100 биаллельных маркёров и 17 Y-STR (коротких тандемных повторов). Анализ полученных данных выявил различия в происхождении и популяционной истории народов Алтая: северные алтайские этносы оказались близки к северным народам, говорящим на енисейских, угорских и самоедских языках, тогда как южные алтайцы приближены к другим тюркоговорящим народам южной Сибири и Центральной Азии. Кроме того, подробный анализ Y-гаплогруппы Q привёл к пересмотру филогении этой ветви, что более наглядно демонстрирует родство популяций Старого и Нового Света и доказывает наличие общего предка у южных алтайцев и американских индейцев. Эти результаты существенно углубляют наше понимание истории заселения Сибири и Америки. (Опубликовано: Am J Hum Genet 2012. 90(3):573).
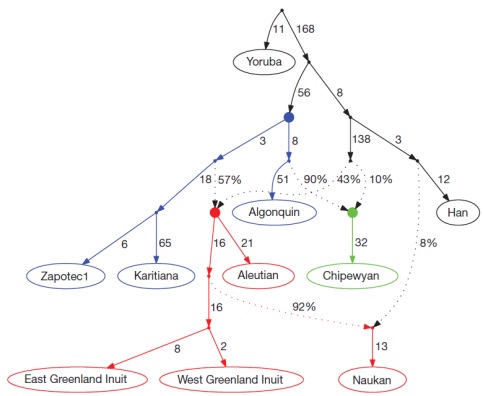
Заселение Америки является предметом обширных генетических, археологических и лингвистических исследований. Однако главные вопросы в этой области остаются до конца не разрешёнными. Один из таких спорных вопросов состоит в том, произошло ли переселение людей из Сибири в Америку за одну волну миграции, или таких волн было несколько. Не совсем ясны и пути расселения людей внутри Американского континента. Чтобы ответить на эти вопросы на уровне современных методов, были проанализированы данные, полученные при генотипировании 52 американских и 17 сибирских популяций по 364 470 однонуклеотидным полиморфным сайтам (SNP). Показано, что коренное население Америки сформировалось на основе по крайней мере трёх потоков генов из Азии. Большая часть населения Америки происходит от одной предковой популяции, которую можно условно назвать «Первые американцы». Однако народы Арктики, говорящие на эскимосско-алеутских языках, унаследовали почти половину генов от более поздних переселенцев из Азии (вторая волна миграции), а у представителей народа чипевьян в Канаде, говорящих на языках на-дене, приблизительно одна десятая часть генов появилась в результате третьей волны миграции. Также показано, что первичное заселение Америки происходило в южном направлении вдоль побережья. Последовавшее за этим разделение популяций сохранялось довольно долго, особенно в Южной Америке. Исключением являются народы, живущие по обе стороны Панамского перешейка и говорящие на чибчанских языках, генофонд которых объединяет компоненты из Северной и Южной Америки. (Опубликовано: Nature 2012. 488(7411):370-4).
Рисунок 5. Генное дерево, представляющее схему переноса генетической информации из Азии в Америку. Пунктирные линии обозначают вторичное смешение ранее разделённых популяций (указаны соответствующие генетические пропорции). Сплошными кругами представлены гипотетические предковые популяции. Синим цветом обозначена первая волна миграции, давшая начало большинству народов Америки, красным – предки алеутов и эскимосов (инуитов), зелёным – предки чипевьян. Также показана обратная миграция эскимосско-алеутских племён через Берингов пролив, приведшая к появлению генов первых американцев у народов северо-восточной Азии (чукчей, науканов).
Были изучены 701 линия митохондриальной ДНК (мтДНК) и 318 Y-хромосом пяти коренных этносов республики Саха (якуты, эвенки, эвены, юкагиры и долганы). Также был проведён анализ более 500 000 аутосомальных однонуклеотидных полиморфных сайтов (SNP) у 758 человек, представляющих 55 этносов, включая 40 ранее не исследованных образцов из Сибири. Концевые ветви восточноазиатских митохондриальных гаплогрупп C и D, а также Y-гаплогруппы N1c, N1b и C3, которые составляют основу генофонда народов республики Саха, соединяют Якутию с южной Сибирью. Анализ данных по аутосомальным SNP подтверждает генетическую связь между народами Саха и южной Сибири. Материнские линии D5a2a2, C4a1c, C4a2, C5b1b и ветвь Y-гаплогруппы N1c с присущим только якутам набором коротких тандемных повторов (STR) могли возникнуть в результате миграции предков современных якутов, тогда как отцовская линия C3c, скорее всего, появилась у населения Саха в результате экспансии тунгусских народов. Гаплогруппы мтДНК Z1a1b и Z1a3, присутствующие у юкагиров, эвенов и долган, указывают на другие, возможно, более древние миграционные события. Анализ как гаплоидных локусов (мтДНК и Y-хромосомы), так и аутосомальных SNP показал, что у народов Саха и у народов северо-восточной Сибири мало общих генетических компонент. Большая часть западно-евразийских материнских и отцовских гаплогрупп у народов Саха, по-видимому, происходит от недавней метисации с восточными европейцами, однако гаплогруппы мтДНК H8, H20a и HV1a1a, а также Y-гаплогруппа J, вероятно, являются следами древнего переноса генов из западной Евразии через Центральную Азию и южную Сибирь. (Опубликовано: BMC Evol Biol 2013. 13:127).
5. Задачи, планируемые на перспективу а) Комплексное исследование популяций тундровых ненцев арктического Севера с целью выявления их демографического потенциала в условиях социальных и техногенных преобразований; оценки роли факторов микроэволюции в поддержании популяционной структуры; сбор образцов фракций крови для дальнейших исследований. б) Изучение процессов адаптации человека к условиям Крайнего Севера на молекулярно-генетическом, биохимическом и физико-химическом уровнях (изучение уровня гормонов и липидов, а также структуры гемоглобина и свойств эритроцитарных мембран с учетом типа питания ненцев выполняется совместно с ФГБУ «НИИ биохимии» СО РАМН) в) Молекулярно-генетические исследования Y-хромосомы, митохондриальной ДНК, а также аутосомальных локусов в популяциях коренных этносов с помощью современных методов экзомного секвенирования и полногеномного сканирования для уточнения эволюционных этапов заселения Евразии и Америки, а также выявления локусов, ответственных за процессы адаптации к экстремальному климату (совместно с зарубежными учёными). г) Исследования генов ферментов биотрансформации ксенобиотиков, участвующих в канцерогенезе - глютатион-S-трансферраз (GSTs), цитохромов (CYP), а также генов систем свёртывания крови (VCORC1, FV, FII), у коренных жителей Сибири в сравнении с европейцами в контексте выявления адаптивных паттернов в зависимости от типа питания и региона проживания (совместно с ФГБУ «Институт химической биологии и фундаментальной медицины»). д) Изучение разнообразия молекулярных вариантов гепатита В в популяциях Сибири и Алтая в прикладном аспекте для разработки оригинальных отечественных тест-систем для диагностики (совместно с ЗАО «Вектор-Бест») Вавилова Наталья Александровна [старший лаборант] Личман Дарья Вениаминовна [ведущий инженер] Молетотова Наталья Анатольевна [старший лаборант] Табиханова Людмила Эдмундовна [младший научный сотрудник] Тийс Роза Павловна [младший научный сотрудник] Чуркина Татьяна Валерьевна [младший научный сотрудник] Бывшие сотрудникиБурлакова Нина АндреевнаГончарова Маргарита Викторовна Ельцов Николай Павлович Зубков Евгений Андреевич Карпелева Наталья Валерьевна Никитина Ольга Николаевна Осипова Людмила Павловна Табиханова Зоя Евгеньевна Выберите слайдером нужный промежуток, и список ниже будет содержать записи только нужного периода: 05 06 07 08 09 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 Публикации Монографии Конференции Гранты
|

| 2023 | ПОВЫШЕННЫЕ ЧАСТОТЫ АЛЛЕЛЕЙ ‒174G И ‒572C ГЕНА IL6 В ПОПУЛЯЦИЯХ КОРЕННЫХ НАРОДОВ СИБИРИ ПО СРАВНЕНИЮ С РУССКИМИ Л. Э. Табиханова, Л. П. Осипова, Т. В. Чуркина, С. С. Ковалев, М. Л. Филипенко, Е. Н. Воронина Молекулярная биология, 2023, Vol. 57, No. 2, pp. 350–359. |
| Increased Frequencies of the ‒174G and ‒572C IL6 Alleles
in Populations of Indigenous Peoples
of Siberia Compared to Russians L. E. Tabikhanova, L. P. Osipova, T. V. Churkina, S. S. Kovalev, M. L. Filipenko, and E. N. Voronina Molecular Biology, 2023, Vol. 57, No. 2, pp. 329–337 |
|
| 2022 | Полиморфизм гена TCF7L2
в популяциях пяти этносов Сибири Л.Э. Табиханова, Л.П. Осипова, Т.В. Чуркина, Е.Н. Воронина, М.Л. Филипенко Vavilov journal of genetics and breeding, 2022, 26(2):188-195 |
| Интерлейкин-6: его роль в организме, генетический полиморфизм и значение при некоторых заболеваниях (литературный обзор) Тийс Роза Павловна, Осипова Людмила Павловна Медицинская генетика, 2022, том 21, № 1 |
|
| Маркеры вирусного гепатита B в образцах плазмы крови коренного населения Крайнего Севера России. Генотипы ВГВ и субтипы HBsAg 59) Безуглова Л.В., Осипова Л.П., Сергеева Е.И., Делий И.В., Табиханова Л.Э., Нетесов С.В., Нетесова И.Г. Молекулярная генетика, микробиология и вирусология, 2022, 40(3):41‑48 |
|
| The Markers of viral hepatitis B in blood plasma samples of the indigenous population of the Far North of Russia. HBV genotypes and HBsAg subtypes Bezuglova LV, Osipova LP, Sergeeva EI, Deliy IV, Tabikhanova LE, Netesov SV, Netesova IG. MOL GENET MICROBIOL+, 2022, 40(3):41‑48 |
|
| Genetic diversity and possible origins of the hepatitis B virus in Siberian natives Victor Manuylov, Vladimir Chulanov, Ludmila Bezuglova, Elena Chub, Anastasia Karlsen, Karen Kyuregyan, Yulia Ostankova, Alexander Semenov, Ludmila Osipova, Tatjana Tallo, Irina Netesova, Artem Tkachuk, Vladimir Gushchin, Sergey Netesov, Lars O. Magnius, Heléne Norder VIRUSES-BASEL, 2022, 14(11):2465 |
|
| TCF7L2 gene polymorphism
in populations of f ive Siberian ethnic groups L.E. Tabikhanova, L.P. Osipova, T.V. Churkina, E.N. Voronina, M.L. Filipenko Vavilov journal of genetics and breeding, 2022, 26(2):188-195 |
|
| 2021 | Molecular characteristic of treatment failure clinical isolates of Leishmania major Gilda Eslami, Samira Hatefi, Vahid Ramezani, Masoud Tohidfar, Tatyana V. Churkina, Yuriy L. Orlov, Saeedeh Sadat Hosseini, Mohammad Javad Boozhmehrani, Mahmood Vakili PeerJ, 2021, PeerJ 9:e10969 https://doi.org/10.7717/peerj.10969 |
| ПОЛИМОРФИЗМ ВАРИАНТОВ ГЕНА N-АЦЕТИЛТРАНСФЕРАЗЫ 2 (NAT2)
И АНАЛИЗ ГЕННОЙ СЕТИ. Тийс Роза Павловна, Осипова Людмила Павловна,Галиева Эльвира Расимовна, Личман Дарья Вениаминовна, Воронина Елена Николаевна,Мелихова А. В., Орлов Юрий Львович, Филипенко Максим Леонидович Биомедицинская химия, 2021, том 67, вып. 3, с. 213-221. |
|
| 2020 | ОСОБЕННОСТИ ГЕНОФОНДА
ТУВИНСКОГО ЭТНОСА НА ОСНОВЕ
ИЗУЧЕНИЯ АНТИГЕНОВ ГРУПП КРОВИ
В СРАВНИТЕЛЬНОМ АСПЕКТЕ Л.Э. Табиханова, Л.П. Осипова, С.С.Ковалёв, У.Н. Кавай-оол, А.Н. Сарыглар, Т.В. Чуркина, М.Р. Хантемирова Якутский медицинский журнал, 2020, №2, с.84-87 |
| FEATURES OF THE GENE POOL
OF THE TUVINIAN ETHNOS BASED
ON THE STUDY OF BLOOD GROUP ANTIGENS IN COMPARATIVE PERSPECTIVE L.E. Tabikhanova, L.P. Osipova, S.S. Kovalev, U.N. Kavai-ool, A.N. Sariglar, T.V. Churkina, M.R. Khantemirova Якутский медицинский журнал, 2020, №2, с.84-87 |
|
| Genetics researches at the “Centenary of human population genetics” conference and SBB-2019 Tatarinova TV, Tabikhanova LE, Eslami G, Bai H, Orlov YL BMC GENET, 2020, 21(Suppl 1): 109 |
|
| Studying polymorphic variants of the NAT2 gene (NAT2*5 and NAT2*7) in Nenets populations of Northern Siberia Tiis R.P., Osipova L.P., Lichman D.V., Voronina E.N., Filipenko M.L. BMC GENET, 2020, 2020; 21(Suppl 1):115 |
|
| Популяционные частоты полиморфизма C1173T гена VKORC1, определяющего чувствительность к варфарину, у лесных ненцев и нганасан Северной Сибири Тийс Р.П., Осипова Л.П., Воронина Е.Н., Филипенко М.Л. Медицинская генетика, 2020, 19(2): 35-42. |
|
| Результаты испытаний реагентов для иммуноферментного определения субтипа HBsAg и генотипа вируса гепатита B в
образцах плазмы крови человека Безуглова Л.В., Мануйлов В.А., Осипова Л.П., Мосина Я.Д., Порываева В.А., Агафонова О.А., Могильных А.К., Нетесов С.В., Нетесова И.Г. Молекулярная генетика, микробиология и вирусология, 2020, 38(4):188–195. |
|
| Trial Results for ELISA Test Kits for HBsAg Subtype and Hepatitis B Virus Genotype Identification in Human Blood Plasma L. V. Bezuglova, V. A. Manuilov, L. P. Osipova, Ya. D. Mosina, V. A. Poryvaeva, O. A. Agafonova, A. K. Mogilnykh, S. V. Netesov, and I. G. Netesova MOL GENET MICROBIOL+, 2020, Vol. 35, No. 4, pp. 229–236. |
|
| 2019 | Genomic Evidence of Local Adaptation to Climate and Diet in Indigenous Siberians Hallmark B, Karafet TM, Hsieh P, Osipova LP, Watkins JC, Hammer MF MOL BIOL EVOL, 2019, 2019 Feb 1;36(2):315-327 |
| Распределение полиморфных вариантов генов биотрансформации ксенобиотиков GSTM1, GSTТ1 и GSTP1, в популяциях коренных жителей и русских Восточной Сибири Табиханова Л.Э., Осипова Л.П., Воронина Е.Н., Филипенко М.Л. Медицинская генетика, 2019, 18(2):24-34 |
|
| Тестирование безопасности продуктов из генетически модифицированного риса: экспериментальное исследование на крысах Спраг-Доули Ширдели М., Орлов Ю.Л., Ислами Г., Хаджимохаммади Б., Табиханова Л.Э., Ихрампуш М.Х., Резвани М.Э., Фаллахзаде Х., Занди Х., Хоссейни С.С., Ахмадиан С., Мортазави Ш., Фаллахи Р., Асади-Юсефабад С.Л. Генетика, 2019, Т. 55. № 8. С. 912-919. |
|
| Особенности липидного спектра и взаимосвязь отдельных классов липидов с уровнем половых гормонов у мужчин на Азиатском Севере Ким Л.Б., Осипова Л.П., Розуменко А.А., Путятина А.Н., Русских Г.С., Козарук Т.В., Воронина Н.П. Якутский медицинский журнал, 2019, №3(67), 2019. С. 27-31 |
|
| Полиморфизм генов липидного обмена
в некоторых популяциях Южной и Восточной Сибири Л.Э. Табиханова, Л.П. Осипова, Е.Н. Воронина, А.О. Брагин, М.Л. Филипенко Vavilov journal of genetics and breeding, 2019, 23(8):1011-1019 |
|
| Polymorphism of lipid exchange genes
in some populations of South and East Siberia L.E. Tabikhanova, L.P. Osipova, E.N. Voronina, 3, A.O. Bragin, M.L. Filipenko Vavilov journal of genetics and breeding, 2019, 23(8):1011-1019 |
|
| СОВРЕМЕННАЯ ГЕНОМИКА В ИССЛЕДОВАНИЯХ АДАПТАЦИИ К КЛИМАТУ КОРЕННЫХ СИБИРСКИХ ПОПУЛЯЦИЙ Осипова Л.П., Личман Д.В., Табиханова Л.Э., Hallmark B., Karafet T.M., Hsieh P.H., Watkins J.C., Hammer M.F. Российский вестник перинатологии и педиатрии, 2019, Т. 64. № 4. С. 226-227. |
|
| 2018 | Полиморфизм генов CYP1A1 и CYP2D6 в популяциях бурят, телеутов и у русских Восточной Сибири Табиханова Л.Э., Осипова Л.П., Чуркина Т.В., Воронина Е.Н., Филипенко М.Л. Vavilov journal of genetics and breeding, 2018, 22(2):205-211 |
| ПОЛИМОРФИЗМ ГЕНОВ F2, F5 И VKORC1
В ПОПУЛЯЦИЯХ БУРЯТ, ТЕЛЕУТОВ И У РУССКИХ
ЖИТЕЛЕЙ ВОСТОЧНОЙ СИБИРИ Л.Э. Табиханова, Л.П. Осипова, Т.В. Чуркина, Е.Н. Воронина, М.Л. Филипенко Молекулярная медицина, 2018, 16 (3): 31–36 |
|
| Genes reveal traces of common recent demographic history for most of the Uralic-speaking populations Kristiina Tambets, Bayazit Yunusbayev, Georgi Hudjashov, Anne-Mai Ilumäe, Siiri Rootsi, Terhi Honkola, Outi Vesakoski, Quentin Atkinson, Pontus Skoglund, Alena Kushniarevich, Sergey Litvinov, Maere Reidla, Ene Metspalu, Lehti Saag, Timo Rantanen, Monika Karmin, Jüri Parik, Sergey I. Zhadanov, Marina Gubina, Larisa D. Damba, Marina Bermisheva, Tuuli Reisberg, Khadizhat Dibirova, Irina Evseeva, Mari Nelis, Janis Klovins, Andres Metspalu, Tõnu Esko, Oleg Balanovsky, Elena Balanovska, Elza K. Khusnutdinova, Ludmila P. Osipova, Mikhail Voevoda, Richard Villems, Toomas Kivisild, Mait Metspalu GENOME BIOL, 2018, (2018) 19:139 |
|
| Генетические исследования мультифакторных заболеваний: системно-биологические подходы Хантемирова М.Р., Табиханова Л.Э., Орлов Ю.Л., Осипова Л.П. Гены и клетки, 2018, Приложение №1: С. 106-107. |
|
| Siberian genetic diversity reveals complex origins of the
Samoyedic-speaking populations Tatiana M. Karafet, Ludmila P. Osipova, Olga V. Savina, Brian Hallmark, Michael F. Hammer AM J HUM BIOL, 2018, 2018;e23194. |
|
| 2017 | C1473G polymorphism in mouse tryptophan hydroxylase-2 gene in the regulation of the reaction to emotional stress Ekaterina Y. Bazhenova,∗, Daria V. Bazovkina, Elizabeth A. Kulikova, Dariya V. Fursenko, Nikita V. Khotskin, Daria V. Lichman, Alexander V. Kulikov Neuroscience Letters, 2017, Neuroscience Letters 640 (2017) 105–110 |
| Компьютерные средства анализа транскриптомных данных: программный комплекс ExpGene А. М. Спицина, А. О. Брагин, А. И. Дергилев, И. В. Чадаева, Н. Н. Твердохлеб, Э. Р. Галиева, Л. Э. Табиханова, Ю. Л. Орлов Програмные системы: теория и приложения, 2017, 8:2(33), 45–68. |
|
| Анализ геномных полиморфизмов в популяциях с помощью секвенирования на примере выборки монголов Китая Табиханова Л.Э., Чен М., Бай Х., Осипова Л.П., Орлов Ю.Л. Acta Naturae, 2017, 9(1), С. 22 |
|
| Генетический полиморфизм и сопутствующие факторы риска ишемического инсульта в монгольской популяции Китая Чижу У, Хуигуань У, Юрий Л. Орлов, Гегентана Гегентана, Веньян Хуо, Анатолий О. Брагин, Нуомин У, Суялату Суялату, Фукуан Жао, Жиньвен Жао, Людмила Э. Табиханова, Минг Чен, Хайхуа Бай. Vavilov journal of genetics and breeding, 2017, Т.21. №5. С.581-587. |
|
| Exome Sequencing Provides Evidence of Polygenic Adaptation to a Fat-Rich Animal Diet in Indigenous Siberian Populations Hsieh P, Hallmark B, Watkins J, Karafet TM, Osipova LP, Gutenkunst RN, Hammer MF MOL BIOL EVOL, 2017, 34(11):2913-2926. |
|
| Анализ содержания инсулина, кортизола и глюкозы в сыворотке крови поселковых жителей Ямало-Ненецкого автономного округа О.Н. Потеряева, Л.П. Осипова, Г.С. Русских, Н.Г. Биушкина, А.А. Розуменко, Т.В. Чуркина, А.В. Зубова, Т.В. Гольцова, Л.М. Поляков Физиология человека, 2017, том 43, № 6, с. 103–108 |
|
| 2016 | Полиморфизм гена цитохрома Р450 CYP1A1 (ILE462VAL) в популяциях тундровых ненцев Ямало-Ненецкого автономного округа, нганасан Таймыра и русских Сибири. Тийс Р.П., Осипова Л.П., Чуркина Т.В., Табиханова Л.Э., Личман Д.В., Воронина Е.Н., Филипенко М.Л. Vavilov journal of genetics and breeding, 2016, Т. 20. № 1. С. 16-22. |
| Полиморфные варианты гена системы биотрансформации ксенобиотиков GSTP1 у ненцев и нганасан тундры по сравнению с русскими Тийс Р.П., Осипова Л.П., Личман Д.В., Табиханова Л.Э., Чуркина Т.В., Воронина Е.Н., Филипенко М.Л., Кривощеков С.Г. Физиология человека, 2016, Т .42. № 2. С. 112-120. DOI: 10.7868/S0131164616020168 |
|
| Human Y Chromosome Haplogroup N: A Non-trivial Time-Resolved Phylogeography that Cuts across Language Families Ilumäe AM, Reidla M, Chukhryaeva M, Järve M, Post H, Karmin M, Saag L, Agdzhoyan A, Kushniarevich A, Litvinov S, Ekomasova N, Tambets K, Metspalu E, Khusainova R, Yunusbayev B, Khusnutdinova EK, Osipova LP, Fedorova S, Utevska O, Koshel S, Balanovska E, Behar DM, Balanovsky O, Kivisild T, Underhill PA, Villems R, Rootsi S AM J HUM GENET, 2016, 2016 Jul 7;99(1):163-73. doi: 10.1016/j.ajhg.2016.05.025. |
|
| Genomic analyses inform on migration events during the peopling of Eurasia Pagani L, Lawson DJ, Jagoda E, Mörseburg A, Eriksson A, Mitt M, Clemente F, Hudjashov G, DeGiorgio M, Saag L, Wall JD, Cardona A, Mägi R, Sayres MA, Kaewert S, Inchley C, Scheib CL, Järve M, Karmin M, Jacobs GS, Antao T, Iliescu FM, Kushniarevich A, Ayub Q, Tyler-Smith C, Xue Y, Yunusbayev B, Tambets K, Mallick CB, Saag L, Pocheshkhova E, Andriadze G, Muller C, Westaway MC, Lambert DM, Zoraqi G, Turdikulova S, Dalimova D, Sabitov Z, Sultana GN, Lachance J, Tishkoff S, Momynaliev K, Isakova J, Damba LD, Gubina M, Nymadawa P, Evseeva I, Atramentova L, Utevska O, Ricaut FX, Brucato N, Sudoyo H, Letellier T, Cox MP, Barashkov NA, Škaro V, Mulahasanovic L, Primorac D, Sahakyan H, Mormina M, Eichstaedt CA, Lichman DV, Abdullah S, Chaubey G, Wee JT, Mihailov E, Karunas A, Litvinov S, Khusainova R, Ekomasova N, Akhmetova V, Khidiyatova I, Marjanović D, Yepiskoposyan L, Behar DM, Balanovska E, Metspalu A, Derenko M, Malyarchuk B, Voevoda M, Fedorova SA, Osipova LP, Lahr MM, Gerbault P, Leavesley M, Migliano AB, Petraglia M, Balanovsky O, Khusnutdinova EK, Metspalu E, Thomas MG, Manica A, Nielsen R, Villems R, Willerslev E, Kivisild T, Metspalu M NATURE, 2016, 2016 Sep 21;538(7624):238-242. [Epub ahead of print] |
|
| Гормональный статус и липидный спектр у жителей пос. Самбург Ямало-Ненецкого автономного округа Потеряева О.Н., Русских Г.С., Биушкина Н.Г., Розуменко А.А., Чуркина Т.В., Гольцова Т.В., Осипова Л.П., Поляков Л.М. Якутский медицинский журнал, 2016, Т. 54. № 2. С. 35-36. |
|
| Эндокринный статус у женщин – представительниц коренного и пришлого населения Ямало-Ненецкого автономного округа Поляков Л.М., Розуменко А.А., Русских Г.С., Биушкина Н.Г., Потеряева О.Н., Гольцова Т.В., Чуркина Т.В., Осипова Л.П., Siberian scientific medical journal, 2016, № 3 / 2016 / 109-113 УДК 577.2:616-006 |
|
| Микровязкость мембран эритроцитов у жителей пос. Самбург Ямало-Ненецкого автономного округа Мокрушников П.В., Осипова Л.П., Гольцова Т.В. Розуменко А.А. Якутский медицинский журнал, 2016, Т. 54. № 2. С.15-16. |
|
| Полиморфизм гена цитохрома Р450 CYP1A1 (Ile462Val) у потомков смешанных браков тундровых ненцев с русскими Табиханова Л.Э., Осипова Л.П., Чуркина Т.В., Воронина Е.Н., Филипенко М.Л. Научный вестник Ямало-Ненецкого автономного округа, 2016, Выпуск 91(2). С. 71-76. |
|
| Strategies of adaptation under prolonged irradiation vs chronic exposure Korogodina V.L., Grigorkina E.B., Osipova L.P. Genetics, Evolution and Radiation: Crossing Borders, The Interdisciplinary Legacy of Nikolay W. Timofeeff-Ressovsky, 2016, P. 153-167 |
|
| 2015 | Association Analysis of Genetic Variants with Type Diabetes in a Mongolian Population in China. Bai H, Liu H, Suyalatu S, Guo X, Chu S, Chen Y, Lan T, Borjigin B, Orlov YL, Posukh OL, Yang X, Guilan G, Osipova LP, Wu Q, Narisu N. Journal of Diabetes Research, 2015, 2015: 613236. doi: 10.1155/2015/613236. Epub 2015 Jul 28. |
| Genomic evidence for the Pleistocene and recent population history of Native Americans. Raghavan M, Steinrücken M, Harris K, Schiffels S, Rasmussen S, DeGiorgio M, Albrechtsen A, Valdiosera C, Ávila-Arcos MC, Malaspinas AS, Eriksson A, Moltke I, Metspalu M, Homburger JR, Wall J, Cornejo OE, Moreno-Mayar JV, Korneliussen TS, Pierre T, Rasmussen M, Campos PF, Damgaard Pde B, Allentoft ME, Lindo J, Metspalu E, Rodríguez-Varela R, Mansilla J, Henrickson C, Seguin-Orlando A, Malmström H, Stafford T Jr, Shringarpure SS, Moreno-Estrada A, Karmin M, Tambets K, Bergström A, Xue Y, Warmuth V, Friend AD, Singarayer J, Valdes P, Balloux F, Leboreiro I, Vera JL, Rangel-Villalobos H, Pettener D, Luiselli D, Davis LG, Heyer E, Zollikofer CP, Ponce de León MS, Smith CI, Grimes V, Pike KA, Deal M, Fuller BT, Arriaza B, Standen V, Luz MF, Ricaut F, Guidon N, Osipova L, Voevoda MI, Posukh OL, Balanovsky O, Lavryashina M, Bogunov Y, Khusnutdinova E, Gubina M, Balanovska E, Fedorova S, Litvinov S, Malyarchuk B, Derenko M, Mosher MJ, Archer D, Cybulski J, Petzelt B, Mitchell J, Worl R, Norman PJ, Parham P, Kemp BM, Kivisild T, Tyler-Smith C, Sandhu MS, Crawford M, Villems R, Smith DG, Waters MR, Goebel T, Johnson JR, Malhi RS, Jakobsson M, Meltzer DJ, Manica A, Durbin R, Bustamante CD, Song YS, Nielsen R, Willerslev E. SCIENCE, 2015, 349(6250):aab3884. doi: 10.1126/science.aab3884. Epub 2015 Jul 21. |
|
| The C1473G polymorphism in the Tryptophan hydroxylase-2 gene: involvement in ethanol-related behavior in mice Bazovkina D.V., Lichman D.V., Kulikov A.V. Neuroscience Letters, 2015, V.589, P.79-82 |
|
| Association between intelligence quotient and the 5HTTLPR polymorphism of human serotonin transporter coding gene Volf N.V., Sinyakova N.A., Osipova L.P., Kulikov A.V., Belousova L.V. Annals of Neuroscience and Psychology, 2015, V.2, P.6 |
|
| Генетический полиморфизм факторов системы свертывания крови FII и FV в популяциях коренных этносов (тундровых и лесных ненцев, нганасан), русских Северной Сибири и их метисов в связи с риском развития тромбозов. Тийс Р.П., Осипова Л.П., Табиханова Л.Э., Чуркина Т.В., Личман Д.В., Воронина Е.Н., Филипенко М.Л. Медицинская генетика, 2015, № 7, С. 27-31 |
|
| Changes in the Structural Ordering of Hemoglobin under Extreme Conditions of the Arctic Region Kunitsyn V.G., Panin L.E, Osipova L.P., Tabikhanova L.E., Churkina T.V., Rozumenko A.A. Journal of Asian Scientific Research, 2015, 5(1):92-95 |
|
| A recent bottleneck of Y chromosome diversity coincides with a global change in culture Karmin M, Saag L, Vicente M, Wilson Sayres MA, Järve M, Talas UG, Rootsi S, Ilumäe AM, Mägi R, Mitt M, Pagani L, Puurand T, Faltyskova Z, Clemente F, Cardona A, Metspalu E, Sahakyan H, Yunusbayev B, Hudjashov G, DeGiorgio M, Loogväli EL, Eichstaedt C, Eelmets M, Chaubey G, Tambets K, Litvinov S, Mormina M, Xue Y, Ayub Q, Zoraqi G, Korneliussen TS, Akhatova F, Lachance J, Tishkoff S, Momynaliev K, Ricaut FX, Kusuma P, Razafindrazaka H, Pierron D, Cox MP, Sultana GN, Willerslev R, Muller C, Westaway M, Lambert D, Skaro V, Kovačevic L, Turdikulova S, Dalimova D, Khusainova R, Trofimova N, Akhmetova V, Khidiyatova I, Lichman DV, Isakova J, Pocheshkhova E, Sabitov Z, Barashkov NA, Nymadawa P, Mihailov E, Seng JW, Evseeva I, Migliano AB, Abdullah S, Andriadze G, Primorac D, Atramentova L, Utevska O, Yepiskoposyan L, Marjanovic D, Kushniarevich A, Behar DM, Gilissen C, Vissers L, Veltman JA, Balanovska E, Derenko M, Malyarchuk B, Metspalu A, Fedorova S, Eriksson A, Manica A, Mendez FL, Karafet TM, Veeramah KR, Bradman N, Hammer MF, Osipova LP, Balanovsky O, Khusnutdinova EK, Johnsen K, Remm M, Thomas MG, Tyler-Smith C, Underhill PA, Willerslev E, Nielsen R, Metspalu M, Villems R, Kivisild T. GENOME RES, 2015, Apr;25(4):459-66. doi: 10.1101/gr.186684.114. Epub 2015 Mar 13. |
|
| The prevalence of HBsAg subtypes and HBV genotypes in native population groups of Siberia Manuilov V.A., Osipova L.P., Netesova I.G., Chub E.V., Bezuglova L.V., Norder H., Magnius L.O., Netesov S.V. Microbiology and Virology, 2015, V. 30. Issue 1. P. 30-38. |
|
| The Genetic Legacy of the Expansion of Turkic-Speaking Nomads across Eurasia Yunusbayev B, Metspalu M, Metspalu E, Valeev A, Litvinov S, Valiev R, Akhmetova V, Balanovska E, Balanovsky O, Turdikulova S, Dalimova D, Nymadawa P, Bahmanimehr A, Sahakyan H, Tambets K, Fedorova S, Barashkov N, Khidiyatova I, Mihailov E, Khusainova R, Damba L, Derenko M, Malyarchuk B, Osipova L, Voevoda M, Yepiskoposyan L, Kivisild T, Khusnutdinova E, Villems R PLOS GENET, 2015, 11(4):e1005068 |
|
| Распространенность маркеров гельминтозов среди жителей села Гыда Тазовского района в 2014 году Осипова Л.П., Чуркина Т.В., Табиханова Л.Э., Ткаченко Т.Н. Научный вестник Ямало-Ненецкого автономного округа, 2015, № 2. С. 101-111 |
|
| 2014 | Изменение структуры гемоглобина в экстремальных условиях Арктики Куницын В.Г., Панин Л.Е., Осипова Л.П., Табиханова Л.Э., Чуркина Т.В., Розуменко А.А. Вестник уральской медицинской академической науки, 2014, №2. С. 37-39. |
| Гормональный статус, липидный спектр и микровязкость мембран у жительниц Севера Потеряева О.Н., Русских Г.С., Мокрушников П.В., Осипова Л.П., Тузиков Ф.В., Тузикова Н.А. Вестник уральской медицинской академической науки, 2014, №2. С. 149-152. |
|
| Ancient human genomes suggest three ancestral populations for present-day Europeans. Iosif Lazaridis, Nick Patterson, Alissa Mittnik, Gabriel Renaud, Swapan Mallick, Karola Kirsanow, Peter H. Sudmant, Joshua G. Schraiber, Sergi Castellano, Mark Lipson, Bonnie Berger, Christos Economou, Ruth Bollongino, Qiaomei Fu, Kirsten I. Bos, Susanne Nordenfelt, Heng Li, Cesare de Filippo, Kay Prüfer, Susanna Sawyer, Cosimo Posth, Wolfgang Haak, Fredrik Hallgren, Elin Fornander, Nadin Rohland, Dominique Delsate, Michael Francken, Jean-Michel Guinet, Joachim Wahl, George Ayodo, Hamza A. Babiker, Graciela Bailliet, Elena Balanovska, Oleg Balanovsky, Ramiro Barrantes, Gabriel Bedoya, Haim Ben-Ami, Judit Bene, Fouad Berrada, Claudio M. Bravi, Francesca Brisighelli, George B. J. Busby, Francesco Cali, Mikhail Churnosov, David E. C. Cole, Daniel Corach, Larissa Damba, George van Driem, Stanislav Dryomov, Jean-Michel Dugoujon, Sardana A. Fedorova, Irene Gallego Romero, Marina Gubina, Michael Hammer, Brenna M. Henn, Tor Hervig, Ugur Hodoglugil, Aashish R. Jha, Sena Karachanak-Yankova, Rita Khusainova, Elza Khusnutdinova, Rick Kittles, Toomas Kivisild, William Klitz, Vaidutis Kučinskas, Alena Kushniarevich, Leila Laredj, Sergey Litvinov, Theologos Loukidis, Robert W. Mahley, Béla Melegh, Ene Metspalu, Julio Molina, Joanna Mountain, Klemetti Näkkäläjärvi, Desislava Nesheva, Thomas Nyambo, Ludmila Osipova, Jüri Parik, Fedor Platonov, Olga Posukh, Valentino Romano, Francisco Rothhammer, Igor Rudan, Ruslan Ruizbakiev, Hovhannes Sahakyan, Antti Sajantila, Antonio Salas, Elena B. Starikovskaya, Ayele Tarekegn, Draga Toncheva, Shahlo Turdikulova, Ingrida Uktveryte, Olga Utevska, René Vasquez, Mercedes Villena, Mikhail Voevoda, Cheryl A. Winkler, Levon Yepiskoposyan, Pierre Zalloua, Tatijana Zemunik, Alan Cooper, Cristian Capelli, Mark G. Thomas, Andres Ruiz-Linares, Sarah A. Tishkoff, Lalji Singh, Kumarasamy Thangaraj, Richard Villems, David Comas, Rem Sukernik, Mait Metspalu, Matthias Meyer, Evan E. Eichler, Joachim Burger, Montgomery Slatkin, Svante Pääbo, Janet Kelso, David Reich, Johannes Krause. NATURE, 2014, 2014 Sep 18;513(7518):409-13. doi: 10.1038/nature13673. |
|
| Upper Palaeolithic Siberian genome reveals dual ancestry of Native Americans Raghavan M, Skoglund P, Graf KE, Metspalu M, Albrechtsen A, Moltke I, Rasmussen S, Stafford TW Jr, Orlando L, Metspalu E, Karmin M, Tambets K, Rootsi S, Mägi R, Campos PF, Balanovska E, Balanovsky O, Khusnutdinova E, Litvinov S, Osipova LP, Fedorova SA, Voevoda MI, DeGiorgio M, Sicheritz-Ponten T, Brunak S, Demeshchenko S, Kivisild T, Villems R, Nielsen R, Jakobsson M, Willerslev E NATURE, 2014, 2014 Jan 2;505(7481):87-91 |
|
| 2013 | Autosomal and uniparental portraits of the native populations of Sakha (Yakutia): implications for the peopling of Northeast Eurasia Fedorova SA, Reidla M, Metspalu E, Metspalu M, Rootsi S, Tambets K, Trofimova N, Zhadanov SI, Hooshiar Kashani B, Olivieri A, Voevoda MI, Osipova LP, Platonov FA, Tomsky MI, Khusnutdinova EK, Torroni A, Villems R. BMC EVOL BIOL, 2013, 2013 Jun 19;13:127 |
| 2012 | A pharmacological evidence of positive association between mouse intermale aggression and brain serotonin metabolism Kulikov A.V., Osipova (Lichman) D.V., Naumenko V.S., Terenina E., Mormède P., Popova N.K. BEHAV BRAIN RES, 2012, V.233:113-119. |
| Полиморфизм гена VKORC1 (C1173T), определяющего чувствительность индивидов к варфарину, в популяциях коренных этносов селькупов и тундровых ненцев, а также у русских Северной Сибири Корчагина Р.П., Осипова Л.П., Вавилова Н.А., Воронина Е.Н., Филипенко М.Л. Вестник кемеровского государственного университета, 2012, Т. 49. № 1. С. 19-23 |
|
| Анализ генетико-демографической структуры контрастных по условиям обитания популяций Агинского Бурятского округа Табиханова Л.Э., Осипова Л.П. RUSS J GENET+, 2012, Т.48. №12. С. 1418-1426 |
|
| Reconstructing Native American population history David Reich, Nick Patterson, Desmond Campbell, Arti Tandon, Stéphane Mazieres, Nicolas Ray, Maria V. Parra, Winston Rojas, Constanza Duque, Natalia Mesa, Luis F. García, Omar Triana, Silvia Blair, Amanda Maestre, Juan C. Dib, Claudio M. Bravi, Graciela Bailliet, Daniel Corach, Tábita Hünemeier, Maria Cátira Bortolini, Francisco M. Salzano, María Luiza Petzl-Erler, Victor Acuña-Alonzo, Carlos Aguilar-Salinas, Samuel Canizales-Quinteros,Teresa Tusie-Luna, Laura Riba, Maricela Rodriguez-Cruz,Mardia Lopez-Alarcon, Ramon Coral-Vazquez, Thelma Canto-Cetina, Irma Silva-Zolezzi, Juan Carlos Fernandez-Lopez, Aleandra V. Conteraras, Gerardo Jimenez-Sanchez, Maria Jose Gomez-Vazques, Julio Molina, AngelCarracedo, Antonio Salas, Carla Gallo, Giovanni Poletti, David B. Witonsky, Gorka Alkorta-Aranburu, Rem I. Sukernik, Ludmila Osipova, Fedorova S., Vasquez R., Villena M., Labuda D., Barrante R., Excoffier L., Bedoya G., Rothhammer F., Dugoujon J.M., Larrouy G., Schart J., Pauls D., Klitz W., Kidd J., Kidd K., Di Rienzo A., Freimer N.B., Alkes L. Price & Andres Ruiz-Linares NATURE, 2012, V. 488. P. 370-374 |
|
| Mitochondrial DNA and Y Chromosome Variation Provides Evidence for a Recent Common Ancestry between Native Americans and Indigenous Altaians Matthew C. Dulik, Sergey I. Zhadanov, Ludmila P. Osipova, Ayken Askapuli, Lydia Gau, Omer Gokcumen, Samara Rubinstein, Theodore G. Schurr AM J HUM GENET, 2012, V. 90. Issue 2, February 2012, P. 229-246. |
|
| 2011 | Association between tryptophan hydroxylase-2 genotype and the antidepressant effect of citalopram and paroxetine on immobility time in the forced swim test in mice Kulikov A.V., Tikhonova M.A., Osipova (Lichman) D.V., Kulikov V.A., Popova N.K. PHARMACOL BIOCHEM BE, 2011, №4, V.99, P. 683-687. |
| Полиморфизм генов биотрансформации ксенобиотиков GSTM1, GSTT1, CYP2D6,вероятных маркеров риска онкологических заболеваний, в популяциях коренных этносов и русских Северной Сибири Р.П. Корчагина., Л.П. Осипова, Н.А. Вавилова, Н.А. Ермоленко, Е.Н. Воронина, М.Л. Филипенко Vavilov journal of genetics and breeding, 2011, Том 15, № 3, С. 448-461 |
|
| Генетический полиморфизм цитохрома Р450 2C9, участвующего в метаболизме лекарственных препаратов, в популяциях коренных жителей Северной Сибири Корчагина Р.П., Осипова Л.П., Вавилова Н.А., Воронина Е.Н., Филипенко М.Л. Бюллетень СО РАМН, 2011, декабрь 2011 г., № 6, С. |
|
| Y-chromosome variation in Altaian Kazakhs reveals a common paternal gene pool for Kazakhs and the influence of Mongolian expansion Dulik M.C., Osipova L.P., Schurr T.G. PloS One, 2011, March 11, 2011. V. 6. Issue 3: e17548. doi:10.1371/journal.pone.0017548 |
|
| 2010 | О роли селективного сайленсера freud-1 в регуляции экспрессии гена 5-нт1а рецептора мозга В.С. Науменко, Д.В. Личман (Осипова), А.С. Цыбко Molecular Biology, 2010, №5, том 44, с.898 - 903 |
| Distribution of the C1473G polymorphism in tryptophan hydroxylase 2 gene in laboratory and wild mice Osipova D.V., Kulikov A.V., Mekada K., Yoshiki A., Moshkin M.P., Kotenkova E.V., Popova N.K. GENES BRAIN BEHAV, 2010, V.9, P.537-543 |
|
| Adaptation and Radiation Induced Chromosomal Instability Studied by Statistical Modeling Victoria L. Korogodina, Boris V. Florko and Ludmila P. Osipova. The Open Evolution Journal, 2010, V. 4. P. 12-22. |
|
| Ancient human genome sequence of an extinct Palaeo-Eskimo Morten Rasmussen1,2,25, Yingrui Li2,3,25, Stinus Lindgreen1,4,25, Jakob Skou Pedersen4, Anders Albrechtsen4, Ida Moltke4, Mait Metspalu5, Ene Metspalu5, Toomas Kivisild5,6, Ramneek Gupta7, Marcelo Bertalan7, Kasper Nielsen7, M. Thomas P. Gilbert1,2, Yong Wang8, Maanasa Raghavan1,9, Paula F. Campos1, Hanne Munkholm Kamp1,4, Andrew S. Wilson10, Andrew Gledhill10, Silvana Tridico11,12, Michael Bunce12, Eline D. Lorenzen1, Jonas Binladen1, Xiaosen Guo2,3, Jing Zhao2,3, Xiuqing Zhang2,3, Hao Zhang2,3, Zhuo Li2,3, Minfeng Chen2,3, Ludovic Orlando13, Karsten Kristiansen2,3,4, Mads Bak14, Niels Tommerup14, Christian Bendixen15, Tracey L. Pierre16, Bjarne Grønnow17, Morten Meldgaard18, Claus Andreasen19, Sardana A. Fedorova5,20, Ludmila P. Osipova21, Thomas F. G. Higham9, Christopher Bronk Ramsey10, Thomas v. O. Hansen22, Finn C. Nielsen22, Michael H. Crawford23, Søren Brunak7,24, Thomas Sicheritz-Pontén7, Richard Villems5, Rasmus Nielsen4,8, Anders Krogh2,4, Jun Wang2,3,4 & Eske Willerslev1,2 NATURE, 2010, 463(7282), P. 757-62 |
|
| Встречаемость субгенотипов вируса гепатита В и субтипов HBsAg у коренного населения севера и юго-востока Сибири Мануйлов В.А., Осипова Л.П., Нетесова И.Г., Чуб Е.В., Цой Л.В., Дульбеев Р.В., Алексеева Л.Р., Norder H., Magnius L.O., Нетесов С.В. MOL GENET MICROBIOL+, 2010, №4. С. 31-36 |
|
| 2009 | О некоторых особенностях формирования и анализа распределений индивидуумов по числу и частоте встречаемости аберрантных клеток в лимфоцитах крови Флорко Б.В., Осипова Л.П., Корогодина В.Л. Mathematical Biology & Bioinformatics, 2009, Т. 4. № 2. С. 52-65 |
| Haplotypic background of a private allele at high frequency in the Americas Schroeder KB, Jakobsson M, Crawford MH, Schurr TG, Boca SM, Conrad DF, Tito RY, Osipova LP, Tarskaia LA, Zhadanov SI, Wall JD, Pritchard JK, Malhi RS, Smith DG, Rosenberg NA. MOL BIOL EVOL, 2009, 26(5), P. 995-1016 |
|
| Влияние тироксина на поведение мышей с наследственными различиями в предрасположенности к каталепсии Зубков Е.А., Куликов А.В. B EXP BIOL MED+, 2009, № 2, том 147, с. 177-180 |
|
| 2008 | Исследование ограниченных каскадных моделей случайных полей на плоскости Н.П. Ельцов, В. А. Огородников, С. М. Пригарин Сибирский журнал вычислительной математики, 2008, Том 11 с 405-412. |
| Genetic variation in the enigmatic Altaian Kazakhs of South-Central Russia: Insights into Turkic population history Gokcumen O, Dulik MC, Pai AA, Zhadanov SI, Rubinstein S, Osipova LP, Andreenkov OV, Tabikhanova LE, Gubina MA, Labuda D, Schurr TG AM J PHYS ANTHROPOL, 2008, 136(3), P. 278-293 |
|
| Mitochondrial Genome Diversity in Arctic Siberians, with Particular Reference to the Evolutionary History of Beringia and Pleistocenic Peopling of the Americas N.V. Volodko, E.B. Starikovskaya, I.O. Mazunin, N.P. Eltsov, P.V. Naidenko, D.C. Wallace, and R.I.Sukernik AM J HUM GENET, 2008, V. 82. P. 1084-1100. |
|
| Генетико-демографическое изучение популяции селькупов и пришлого населения по Толькинскому сельсовету Красноселькупского района ЯНАО во временном срезе 1967-1970 гг. Осипова Л.П., Табиханова Л.Э., Сенькова Н.А. Научный вестник Ямало-Ненецкого автономного округа, 2008, Выпуск №7 (59). Генетическая демография и биомедицина: результаты исследований коренного населения Красноселькупского района ЯНАО. Часть 2. Салехард. 2008. С.6-38 |
|
| The Russian Old Believers: genetic consequences of their persecution and exile, as shown by mitochondrial DNA evidence Rubinstein S., Dulik M.C., Gokcumen O., Zhadanov S., Osipova L., Cocca M., Mehta N., Gubina M., Posukh O., Schurr T.G. HUM BIOL, 2008, V. 80, N. 3, P. 203-237. |
|
| Генетико-демографическое изучение популяции селькупов и пришлого населения по Толькинскому сельсовету во временном срезе 1988-1990 Осипова Л.П., Чуркина Т.В., Сенькова Н.А. Научный вестник Ямало-Ненецкого автономного округа, 2008, № 7 (59) |
|
| Utilization of a two-standard system in real-time PCR for quantification of gene expression in the brain Naumenko V.S., Lichman (Osipova) D.V., Kostina E.V., Kulikov A.V. J NEUROSCI METH, 2008, №2, V. 170, P.197-203 |
|
| Анализ сенсомоторной функциональной асимметрии головного мозга среди населения Красноселькупского района ЯНАО Сенькова Н.А., Осипова Л.П. Научный вестник Ямало-Ненецкого автономного округа, 2008, Салехард. 2008. Выпуск №7 (59) С. 39-52 |
|
| Некоторые результаты социобиологического обследования учащихся Красноселькупского и Пуровского районов ЯНАО Осипова Л.П., Бурлакова Н.А., Сенькова Н.А. Научный вестник Ямало-Ненецкого автономного округа, 2008, Салехард. 2008. Выпуск № 7 (59) С.106-116 |
|
| 2007 | A counter clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe Siiri Rootsi, Lev A. Zhivotovsky, Marian Baldovic, Manfred Kayser, Ildus A. Kutuev, Rita Khusainova, Marina A. Bermisheva, Marina Gubina, Sardana A. Fedorova, Anne-Mai Ilum, Elza K. Khusnutdinova, Mikhail I. Voevoda, Ludmila P. EUR J HUM GENET, 2007, Feb;15(2):204-11 |
| A private allele ubiquitious in the Americas Schroeder K.B., Schurr T.G., Long J.C., Rosenberg N.A., Crawford M.H., Tarskaia L.A., Osipova L.P., Zhadanov S.I., Smith D.G. BIOL LETT-UK, 2007, v. 3, № 2, pp. 218-223 |
|
| Динамика генетико-демографических процессов в популяциях коренного населения Аксарковской и Харсаимской сельских администраций на 2003 год Осипова Л.П., Табиханова Л.Э., Чуркина Т.В., Сенькова Н.А., Андреенков О.В., Бурлакова Н.А. Научный вестник Ямало-Ненецкого автономного округа, 2007, Вып. N 8(52). Генетическая демография и биомедицина: результаты исследований коренного населения Приуральского и Красноселькупского районов ЯНАО. Салехард. 2007. С. 5-18. |
|
| Популяционно-генетическое исследование селькупской популяции Красноселькупского сельского совета на 1969 и 1990 года Осипова Л.П., Чуркина Т.В., Табиханова Л.Э., Бурлакова Н.А. Научный вестник Ямало-Ненецкого автономного округа, 2007, Выпуск № 8. (52) Генетическая демография и биомедицина: результаты исследований коренного населения Приуральского и Красноселькупского районов ЯНАО. Салехард. 2007. С. 76-119 |
|
| Динамика генетико-демографической структуры популяции селькупов пос. Ратта Красноселкупского района ЯНАО на 1959, 1981 и 2005 года Л.П.Осипова, Т.В.Чуркина, Н.А.Бурлакова, Д.В.Осипова, Т.М.Карафет Научный вестник Ямало-Ненецкого автономного округа, 2007, № 8 (52) |
|
| Beringian standstill and spread of Native American founders Tamm E., Kivisild T., Reidla M., Metspalu M., Smith D.G., Mulligan C.J., Bravi C.M., Rickards O., Martinez-Labarga C., Khusnutdinova E.K., Fedorova S.A., Golubenko M.V., Stepanov V.A., Gubina M.A., Zhadanov S.I., Osipova L.P., Damba L., Voevoda M.I., Dipi PloS One, 2007, 2(9):e829 |
|
| 2006 | Радиоэкологическая обстановка в местах проживания коренного населения Сибири на рубеже XX-XXI веков Щербов Б.Л., Маликова И.Н., Осипова Л.П., Страховенко В.Д., Сухоруков Ф.В. Проблемы биогеохимии и геохимической экологии, 2006, Т. 3, № 3, С. 13-21. |
| The phylogeography of mtDNA variation in Altaian Kazakh populations Gokсumen O, Osipova LP, Zhadanov SI, Andreenkov O, Tabikhanova L, Gubina M, Schurr TG AM J PHYS ANTHROPOL, 2006, Suppl. 42:93 |
|
| Variability of fire behavior, fire effects, and emissions in Scotch pine forests of central Siberia McRae D., Conard S., Ivanova G., Sukhinin A., Baker S., Samsonov Y, Blake T., Ivanov V., Churkina T., Hao W., Kousenogii K., Kovaleva N. MITIG ADAPT STRAT GL, 2006, v. 11, pp.47-74 |
|
| Генетико-демографическая структура популяций коренных народов Сибири в связи с проблемами микроэволюции Гольцова Т.В., Осипова Л.П. Информационный вестник ВОГИС, 2006, Т. 10, № 1, С. 126 - 154 |
 Монографии
Монографии
| 2021 | A Genetic Perspective on Origin and Migration of Samoyedic-speaking Populations from Siberia (edited by María de Lourdes Muñoz-Moreno and Michael H. Crawford) Tatiana M. Karafet, Ludmila P. Osipova, Michael F. Hammer |
| 2010 | Genetic Diversity in Native Siberians: Implications for the Prehistoric Settlement of the Cis-Baikal Region Schurr T.G., Osipova L.P., Zhadanov S.I., Dulik M.C. |
| 2006 | Некоторые актуальные проблемы современной антропологии отв. ред И.И. Гохман, А.В. Громов |
Конференции
| 2024 | ЧАСТОТА АЛЛЕЛЯ -772A ГЕНА FUT2, АССОЦИИРОВАННОГО С НЕВОСПРИИМЧИВОСТЬЮ К HELICOBACTER PYLORI, В ВЫБОРКАХ ЯКУТОВ И РУССКИХ ВОСТОЧНОЙ СИБИРИ Табиханова Л.Э., Осипова Л.П., Чуркина Т.В., Личман Д.В., Воронина Е.Н., Филипенко М.Л. Международный Конгресс "VIII Съезд Вавиловского общества генетиков и селекционеров, посвященный 300-летию российской науки и высшей школы" |
| FUT2 Gene Polymorphism in samples of Tuvinians and Russians of Eastern Siberia Tabikhanova L. E., Osipova L. P., Churkina T. V., Lichman D. V., Voronina E. N., Filipenko M. L. 14-я Международная мультиконференция «Биоинформатика регуляции и структуры геномов / системная биология» / 14th International Multiconference on “Bioinformatics of Genome Regulation and Structure/Systems Biology”, BGRS/SB-2024. |
|
| Genotyping of the e2, e3, e4 alleles of the APOΕ gene in sam-ples of indigenous populations of Eastern Siberia. TiisR., TabikhanovaL., LichmanD., VoroninaE., OsipovaL., FilipenkoM. 14-я Международная мультиконференция «Биоинформатика регуляции и структуры геномов / системная биология» / 14th International Multiconference on “Bioinformatics of Genome Regulation and Structure/Systems Biology”, BGRS/SB-2024. |
|
| 2022 | Polymorphism rs1800795 of the IL6 gene in samples
of Siberian indigenous ethnic groups Tabikhanova L.E.1*, Osipova L.P.1, Churkina T.V.1, Voronina E.N.2, Filipenko M.L.2 13th International Multiconference “Bioinformatics of Genome Regulation and Structure/Systems Biology” |
| Distribution of total cholesterol and glucose parameters
in a sample of Tuvans in Kyzyl Kovalev S.1*, Churkina T.1, Tabikhanova L.1, Hantemirova M.1, Kavai-Ool U.2, Saryglar A.1, Osipova L.1 13th International Multiconference “Bioinformatics of Genome Regulation and Structure/Systems Biology” |
|
| Полиморфизм rs1800795 гена IL6 в выборках коренных народов Сибири Табиханова Л.Э., Осипова Л.П., Чуркина Т.В., Ковалев С.С., Филипенко М.Л., Воронина Е.Н. Научная конференция «Генетические процессы в популяциях» с международным участием, посвященная 50-летнему юбилею лаборатории популяционной генетики им. Ю.П. Алтухова ИОГен РАН и 85-летию со дня рождения академика Юрия Петровича Алтухова |
|
| Некоторые результаты анализа репродуктивного поведения женщин Тазовского района Ямало-Ненецкого АО Осипова Л.П., Табиханова Л.Э., Чуркина Т.В. Научная конференция «Генетические процессы в популяциях» с международным участием, посвященная 50-летнему юбилею лаборатории популяционной генетики им. Ю.П. Алтухова ИОГен РАН и 85-летию со дня рождения академика Юрия Петровича Алтухова. |
|
| 2021 | ЧАСТОТЫ ВАРИАНТА TCF7L2 103894T, АССОЦИИРОВАННОГО С САХАРНЫМ ДИАБЕТОМ 2 ТИПА, У ЯКУТОВ И РУССКИХ ВОСТОЧНОЙ СИБИРИ Табиханова Л.Э. *, Осипова Л.П., Чуркина Т.В., Воронина Е.Н. Сахарный диабет – 2021: от мониторинга к управлению. |
| 2020 | The rs12255372 and rs7903146 polymorphisms of the
TCF7L2 gene among Buryats and Russians of Eastern Siberia Tabikhanova L.E., Osipova L.P., Churkina T.V., Lichman D.V., Voronina E.N., Filipenko M.L. Bioinformatics of Genome Regulation and Structure/ Systems BiologyThe 12th International Multiconference |
| 2019 | Этнические аспекты в распределении частот генов липидного обмена среди бурят и русских Восточной Сибири Табиханова Л.Э., Осипова Л.П., Воронина Е.Н., Филипенко М.Л. Международный конгресс "VII съезд Вавиловского общества генетиков и селекционеров, посвященный 100-летию кафедры генетики СПБГУ, и ассоциированные симпозиумы" |
| ПОЛИМОРФИЗМ ГЕНА FTO C83401A, АССОЦИИРОВАННОГО С МЕТАБОЛИЧЕСКИМ СИНДРОМОМ, В НЕКОТОРЫХ ПОПУЛЯЦИЯХ ВОСТОЧНОЙ СИБИРИ Л.Э. Табиханова, Л.П. Осипова, Е.Н. Воронина, Т.В. Чуркина, Д.В.Личман, А.О. Брагин, М.Л. Филипенко V Международная научная конференция "Биоразнообразие и сохранение генофонда флоры, фауны и народонаселения Центрально-Азиатского региона" |
|
| Исследование генетических детерминант мультифакторных заболеваний у коренных народов Сибири Табиханова Л.Э. IV Межрегиональный фестиваль "Молодой профессионал Сибири" |
|
| Исследование мультифакторных заболеваний на примере предрасположенности к алкоголизму: молекулярная генетика и биофизика Осипова Л.П., Хантемирова М.Р., Галиева Э.Р., Личман Д.В., Бабенко Р.О., Ковалев С.С., Табиханова Л.Э., Орлов Ю.Л. VI Съезд биофизиков России |
|
| Context complexity of sites containing single nucleotide polymorphisms in human genome Luzin A.N., Dergilev A.I., Tabikhanova Z.E., Safronova N.S., Orlov Y.L. Systems Biology and Bioinformatics (SBB-2019) The Eleventh International Young Scientists School. |
|
| 2018 | Metabolic syndrome associated FTO gene polymorphism in populations of Buryats and Russians of Eastern Siberia L.E. Tabikhanova, L.P. Osipova, E.N. Voronina, M.L. Filipenko BGRS\SB-2018. The 11th International Conference. Systems Biology and Biomedicine (SBioMed-2018) : Symposium |
| Polymorphism CYP2D6 gene for xenobiotic biotransformation in populations of Buryats and Russians of Eastern Siberia L.E. Tabikhanova, L.P. Osipova, E.N. Voronina, M.L. Filipenko. BGRS\SB-2018. The 11th International Conference. Integrative Bioinformatics and Systems Biology (WIBSB-2018): First Sino-Russian Workshop |
|
| Computer genomics of regulatory single nucleotide polymorphisms in neurodegenerative diseases based on metabolic pathways models Shanmughavel P., Sathishkumar R., Ponomarenko M.P., Khantemirova M.R., Tabikhanova L.E., Orlov Y.L. Integrative Bioinformatics and Systems Biology (WIBSB-2018): First Sino-Russian Workshop |
|
| 2017 | ПОЛИМОРФИЗМ ГЕНОВ GSTM1 и GSTT1 В ПОПУЛЯЦИЯХ БУРЯТ, ТЕЛЕУТОВ И У РУССКИХ ВОСТОЧНОЙ СИБИРИ Л.Э. Табиханова, Л.П. Осипова, Т.В. Чуркина, Е.Н. Воронина, М.Л. Филипенко Беляевские чтения : Международная конференция, посвященная 100-летию со дня рождения академика АН СССР Д.К. Беляева |
| Полиморфизм генов-кандидатов формирования алкогольной зависимости в популяциях коренных этносов Сибири Хантемирова М. Р. Осипова Л. П. Воронина Е.Н. Филипенко М.Л. Науменко В. С. Базовкина Д. В. БЕЛЯЕВСКИЕ ЧТЕНИЯ, Международная конференция, посвященная 100-летию со дня рождения академика АН СССР Дмитрия Константиновича Беляева |
|
| 2016 | Полиморфизм гена цитохрома Р450 CYP1A1 (Ile462Val) у потомков смешанных браков тундровых ненцев с русскими Табиханова Л.Э., Осипова Л.П., Чуркина Т.В., Воронина Е.Н., Филипенко М.Л. Биомедицина-2016 |
| The ILE462VAL polymorphism of the cytochrome P450 CYP1A1 gene among Eastern Buryats compared with Russians in Trans Baikal Area L.E. Tabikhanova, L.P. Osipova, T.V. Churkina, E.N. Voronina, M.L. Filipenko BGRS-2016 and Structure\ Systems Biology — BGRS\SB-2016 Symposium Systems Biology and Biomedicine (SBioMed-2016) |
|
| The genetic polymorphism of glutathione S-transferase P1 (GSTP1) among Buryats, Teleuts and Russians L.E. Tabikhanova, L.P. Osipova, T.V. Churkina, H. Bai, E.N. Voronina, M.L. Filipenko BGRS\SB-2016, SBioMed-2016 and Structure\ Systems Biology — BGRS\SB-2016 Symposium Systems Biology and Biomedicine (SBioMed-2016) |
|
| Полиморфизм гена VKORC1 (C1173T) у потомков смешанных браков тундровых тундровых ненцев с русскими Табиханова Л.Э., Осипова Л.П., Чуркина Т.В., Воронина Е.Н., Филипенко М.Л. 50 лет ВОГиС: успехи и перспективы |
|
| 2012 | МОЛЕКУЛЯРНАЯ ДИАГНОСТИКА ДЕЛЕЦИЙ В ГЕНАХ ГЛУТАТИОН-S-ТРАНСФЕРАЗ GSTM1 И GSTT1 СРЕДИ ЛЕСНЫХ НЕНЦЕВ (ДЕТЕЙ И ВЗРОСЛЫХ) Корчагина Р.П., Осипова Л.П., Ермоленко Н.А, Филипенко М.Л. XI Российский конгресс инновационных технологий в педиатрии и детской хирургии |
| 2011 | ГЕНЕТИЧЕСКИЙ ПОЛИМОРФИЗМ ЦИТОХРОМА Р450 2С9, ОТВЕТСТВЕННОГО ЗА МЕТАБОЛИЗМ ВАРФАРИНА, В ПОПУЛЯЦИЯХ ЛЕСНЫХ И ТУНДРОВЫХ НЕНЦЕВ Корчагина Р.П., Осипова Л.П., Воронина Е.Н., Филипенко М.Л. II международная научно-практическая конференция «Постгеномные методы анализа в биологии, лабораторной и клинической медицине: геномика, протеомика, биоинформатика» |
| О генетических основах устойчивости человека к онкологическим заболеваниям по результатам изучения глутатион-S-трансфераз Осипова Л.П., Корчагина Р.П. Актуальные вопросы генодиагностики в современной медицине. Семинар мастер-класс |
|
| Распределение частот полиморфных вариантов генов CYP2D6, GSTM1, GSTT1 - вероятных маркеров предрасположенности к развитию онкопатологий в популяции нганасан Таймыра Корчагина Р.П., Осипова Л.П., Воронина Е.Н., Ермоленко Н.А., Филипенко М.Л. II Всероссийская научно-практическая конференция с международным участием "Медико-биологические аспекты мультифакториальной патологии" |
|
| 2010 | Распределение полиморфных вариантов генов CYP2C9, CYP4F2, VKORC1, ответственных за метаболизм антикоагулянтов в популяции тундровых ненцев Корчагина Р.П., Осипова Л.П., Сенькова Н.А., Воронина Е.Н., Филипенко М.Л. Материалы девятого Российского конгресса "Инновационные технологии в педиатрии и детской хирургии". 19-21 октября 2010 г, Москва |
| Insight into population history, evolution, and demographic events in Northwestern Siberia: identity and interaction Karafet T.M., Osipova L.P., Kuzmin Y.V., Hammer M.F. 7th International conference on bioinformatics of genome regulation and structure\system biology (BGRS/SB'20010). June 20-27, 2010, Novosibirsk |
|
| De novo mutations in Y-chromosome STR loci revealed in paternal leneages of Siberian tundra Nentsi population Osipova L.P., Karafet T.M., Hammer M. Crimean Meeting: Third International Conference, Dedicated N.V. Timofeeff-Ressovsky "Modern Problems of Genetics, Radiobiology, Radioecology and Evolution"; Third Readings after V.I. Korogodin and V.A. Shevchenko; NATO Advanced Research Workshop "Radiobio |
|
| Use of ancient DNA genomic sequencing in assessment of evolutionary changes resulting from human adaptation to North Osipova L.P. Crimean Meeting: Third International Conference, Dedicated N.V. Timofeeff-Ressovsky "Modern Problems of Genetics, Radiobiology, Radioecology and Evolution"; Third Readings after V.I. Korogodin and V.A. Shevchenko; NATO Advanced Research Workshop "Radiobio |
|
| К вопросу о генетических основах психофизиологии шаманов Осипова Л.П., Сенькова Н.А. Международный интердисциплинарный научный симпозиум "Психофизиология и социальная адаптация (нео)шаманов в прошлом и настоящем", Республика Бурятия, Тункинский национальный парк, 2-9 августа 2010 |
|
| Сохранение традиционного образа жизни как основа сохранения генофонда тундровых ненцев Осипова Л.П. Научно-практическая конференция "Обдория: история, культура, современность", г. Салехард, 22 ноября 2010 г. |
|
| Association between the serotonin-related gene polymorphisms and hereditary aggressiveness Kulikov A.V., Bazovkina D.V., Osipova (Lichman) D.V., Naumenko V.S., Popova N.K., Rick D'Earth, Simon Turner, Elena Terenina, Pierre Mormede. the Joint Russian-French seminar "Genomics, Bioinformatics and Life Science Modeling" 18-21 June, Novosibirsk |
|
| New mouse strains for behavioral and psychopharmacological genetics Kulikov A.V., Bazovkina D.V. Osipova D.V., Tikhonova M.A., Kondaurova E.M., Popova N.K 7th International Conference on Bioinformatics of Genome Regulation and Structure\ Systems Biology |
|
| Study of polymorhisms in glutathione-S-transferase M1 and T1 (GSTM1 and GSTT1) genes in Selkups and Tundra Nenets Korchagina R.P., Osipova L.P., Senkova N.A., Kostrykina N.A., Philipenko M.L. The seventh international conference on bioinformatics of genome regulation and structure\system biology |
|
| 2009 | Генетическое разнообразие генофондов коренных народов Сибири Осипова Л.П. Международная конференция "Хромосома 2009", Новосибирск, 31 августа-6 сентября |
| Оценка отдаленных последствий воздействия радиационных факторов от СИП на население Иркутской области (цитогенетический анализ) Осипова Л.П., Сенькова Н.А, Матвеева В.Г., Гайнер Т.А., Медведев В.И. Международная научно-практическая конференция "Семипалатинский испытательный полигон спустя 60 лет". Новосибирск, 22 октября 2009 г. |
|
| Роль цитогенетических исследований в предупреждении рождения детей-инвалидов в отягощенных семьях коренных народностей Севера Осипова Л.П., Сенькова Н.А, Гайнер Т.А. Научная программа VIII Российского конгресса "Современные технологии в педиатрии и детской хирургии". Москва, 20-22 октября 2009 г. |
|
| Радиационное воздействие Семипалатинского ядерного полигона на Южную Сибирь В.И. Медведев, Л.Г. Коршунов, Б.П. Черняго, В.В. Коваленко, Н.И. Маторова, А.Н. Карчевский, Л.П. Осипова, М.П. Кельман Радиоактивность и радиоактивные элементы в среде обитания человека. Материалы III Международной конференции, г. Томск, 23–27 июня 2009 г. |
|
| 2008 | The new algorithm for phylogenetic reconstruction of non-recombining DNA sequences N.P. Eltsov, N.V. Volodko. 6th international conference on bioinformatics of genome regulation and structure BGRS’2008, Novosibirsk, June 22-28 |
| Early Siberian Maternal Lineages in the Tubalar of Northeastern Altai Inferred from High-Resolution Mitochondrial DNA Analysis R. Sukernik, I. Mazunin, E. Starikovskaya, N. Volodko, N. Eltsov. Abstract 2536. Presented at the annual meeting of The American Society of Human Genetics, November 11-15, 2008, Philadelphia, Pennsylvania |
|
| New tool (mtPHYL) proposed for phylogenetic analysis of human complete mitochondrial genomes N. Eltsov, N. Volodko, E. Starikovskaya, R. Sukernik Abstract 2536. Presented at the annual meeting of The American Society of Human Genetics, November 11-15, 2008, Philadelphia, Pennsylvania |
|
| Selection Left Distinctive Signatures At The Human Mitochondrial Genome Level in Siberia/Beringia N.V. Volodko, N.P. Eltsov, P.V. Naidenko, E.B. Starikovskaya, R.I. Sukernik. The XX International Congress of Genetics. Berlin, Germany, July 12 - 17, 2008 |
|
| mtPHYL, a new software tool to analyze human complete mitochondrial genomes N.P. Eltsov and N.V. Volodko. The XX International Congress of Genetics. Berlin, Germany, July 12 - 17, 2008 |
|
| Цитогенетическая оценка отдаленных последствий воздействия факторов радиационной природы на население пос. Малое Голоустное Иркутской области Осипова Л.П., Сенькова Н.А., Гайнер Т.А., Матвеева В.Г. Материалы Российской научной конференции "Медико-биологические проблемы токсикологии и радиологии". Санкт-Петербург, 29-30мая 2008 г. Вестник Российской военно-медицинской академии. |
|
| Параметры репродукции и размер семьи в связи с сохранностью традиционного образа жизни в популяциях народов Севера Осипова Л.П., Табиханова Л.Э., Сенькова Н.А., Чуркина Т.В. VII Российский конгресс "Современные технологии в педиатрии и детской хирургии". |
|
| Традиционный образ жизни как важнейший фактор сохранения репродуктивного поведения и демографических характеристик коренного населения ЯНАО Осипова Л.П. Программа научно-практической конференции "Обдория: история, культура, современность". Садехард, 3 декабря 2008 г. |
|
| mtPhyl – новый инструмент для анализа полных последовательностей митохондриальной ДНК Н.П. Ельцов, Н.В. Володько Технологии Microsoft в теории и практике программирования. Новосибирск. 1-2 марта, 2008 |
|
| Сравнительная характеристика структуры семьи и репродуктивного поведения женщин в поселках с разным образом жизни (Агинский Бурятский автономный округ) Табиханова Л.Э., Осипова Л.П., Леган М.В. VII Российский конгресс "Современные технологии в педиатрии и детской хирургии" |
|
| Цитогенетическая оценка отдаленных последствий воздействия факторов радиационной природы на население пос. Малое Голоустное Иркутской области Осипова Л.П., Сенькова Н.А., Гайнер Т.А., Матвеева В.Г. Медико-биологические проблемы токсикологии и радиологии |
|
| Распределение лиц по частоте встречаемости аберрантных клеток в лимфоцитах крови в выборках жителей Новосибирска, Иркутской и Тюменской областей Антонова Е., Осипова Л.П., Сенькова Н.А., Флорко Б.В., Корогодина В.Л. Медико-биологические проблемы токсикологии и радиологии |
|
| Сравнение структуры распределения лиц до 18 лет и старше по частоте встречаемости аберрантных клеток в лимфоцитах крови в выборках жителей пос. Малое Голоустное и Листвянка Иркутской области Антонова Е., Осипова Л.П., Сенькова Н.А., Флорко Б.В., Корогодина В.Л. Медико-биологические проблемы токсикологии и радиологии |
|
| Статистическое исследование цитогенетических эффектов. Оценка риска хромосомной нестабильности в популяциях, испытывающих влияние малых доз радиации Корогодина В.Л., Флорко Б.Ф., Осипова Л.П., Сенькова Н.А., Антонова Е.С. Радиоэкология: итоги, современное состояние и перспективы. |
|
| 2007 | Динамика показателей репродукции и индекса потенциального отбора Кроу в популяциях коренного населения Сибири в период перехода на оседлость Гольцова Т.В., Осипова Л.П. 7-й Конгресс этнографов и антропологов России: доклады и выступления. Саранск, 9-14 июля 2007 г. Саранск |
| The Phylogeography of Haplogroup N1a Gokcumen O., Dulik M., Zhadanov S.I., Osipova L.P., Schurr T.G. The American Association of Physical Anthropologists Meeting, Philadelphia, March, 2007 |
|
| Further studies on Leber’s hereditary optic neuropathy (LHON) in Russia/Siberia Volodko N, Starikovskaya E, Naidenko P, Eltsov N, Sukernik R Еhe annual meeting of The American Society of Human Genetics, October 23-27, 2007, San Diego, California |
|
| Различная встречаемость субгенотипов вируса гепатита B и субтипов HBsAg у коренного населения Сибири В.А. Мануйлов, Е.В. Чуб, И.Г. Нетесова, Л.П.Осипова, Л.В. Цой, H. Norder, L.O. Magnius, С.В.Нетесов. Материалы VII Российской научно-практической конференции c международным участием <<Вирусные гепатиты - эпидемиология, диагностика, лечение и профилактика>>. Москва, 29-31 мая 2007 г. |
|
| Особенности трансформации генетической структуры сибирских популяций в условиях перехода кочевых народов на оседлость Гольцова Т.В., Осипова Л.П. Проблемы биологической и культурной адаптации человеческих популяций, научная конференция Октябрь 2007, Санкт-Петербург, Музей антропологии и этнографии РАН (Кунсткамера) |
|
| 2006 | Изучение полиморфизмов DEL425 и 3’UTR (TC→CA) гена C-FMS в популяциях коренных народов Западной Сибири Сенькова Н.А., Осипова Л.П., Ромащенко А.Г. 12 Международный конгресс по приполярной медицине (Новосибирск, 12-16 июня 2006 г.) |
| Comparative analysis of two approaches in stochastic simulation of polarized radiation transfer Eltsov N.P., Ogorodnikov V.A., Prigarin S.M. Atmospheric and Ocean Optics. Atmospheric Physics. The XIII international symposium. July 2-6, Tomsk |
|
| Monte Carlo Simulation of Angular Characteristics for Polarized Radiation in Water-Drop and Crystal Clouds Prigarin S.M., Eltsov N.P., Fedchenko N.V. Atmospheric and Ocean Optics. Atmospheric Physics. The XIII international symposium. July 2-6, Tomsk |
|
| Genetic diversity of hepatitis B virus among aboriginal populations of Siberia V.A. Manuilov, E.V. Chub, I.G. Netesova, L.P. Osipova, L.V. Tsoy, H. Norder, L.O. Magnius, S.V. Netesov International Network Meeting "Nosocomial and iatrogenic viral hepatitis in Russia and in the Baltic – Network supported by the Swedish Institute." Riga, Latvia – September 20, 2006 |
|
| Comparison of two statistic modeling algorithms of polarized radiation transfer Eltsov N.P. Proceedings of young scientist conference. Novosibirsk: Inst. of Comp. Math. and Math. Geoph. 2006 |
|
| The phylogeography of mtDNA variation in Altaian Kazakh populations Gokсumen O, Osipova LP, Zhadanov SI, Andreenkov O, Tabikhanova L, Gubina M, Schurr TG The American Association of Physical Anthropologists Meeting |
|
| Mitochondrial DNA story of Turkic-speaking Altai populations: implication for the prehistory of Siberia Zhadanov SI, Osipova L, Dulik M, Gubina M, Schurr TG The American Association of Physical Anthropologists Meeting, Anchorage, March, 2006 |
|
| Mitochondrial DNA variation in Old Believer and ethnic Russian populations of northern Siberia Rubinstein S, Mehta N, Cocca MF, Zhadanov SI, Osipova LP, Schurr TG The American Association of Physical Anthropologists Meeting, Anchorage, March, 2006 |
|
| Отдаленные эффекты радиационного воздействия на коренное население Севера Осипова Л.П. Биологические эффекты малых доз ионизирующей радиации и радиоактивное загрязнение среды: тезисы докладов Международной конференции (Сыктывкар, Республика Коми, Россия, 28 февраля – 3 марта 2006 г.) |
|
| Генетическая вариабельность вируса гепатита В среди населения Западной Сибири Мануйлов В.А., Нетесова И.Г., Осипова Л.П., Нетесов С.В. Геномные технологии в медицине и медицинское образование на рубеже веков. Республика Казахстан, Алматы. 18-20 мая 2006 г. |
|
| Частота встречаемости серологических маркеров инфекций гепатитов В и С у населения Туруханского района Красноярского края Нетесова И.Г., Посух О.Л., Осипова Л.П., Мануйлов В.А., Нетесов С.В. Материалы 13 Международного конгресса по приполярной медицине (Новосибирск, 12-16 июня 2006 г.) |
|
| Сохранение генофондов коренных малочисленных народов в условиях промышленного освоения Севера Осипова Л.П. Тезисы докладов участников совместного заседания Президиума Уральского Отделения РАН и Администрации Ямало-Ненецкого АО. 20-23 июня 2006г., г.Салехард. |
|
| Влияние полиморфизма в гене ТПГ2 на поведение самцов мышей Тибейкина М.А., Куликов А.В., Осипова (Личман) Д.В., Науменко В.С., Попова Н.К. Российская конференция молодых ученых |
|
| The demographic database creation and analysis of the demogenetic structure of contemporary Kets population. Посух Ольга Леонидовна 13 International Congress on Circumpolar Health. Novosibirsk, Russia, June 12-16 |
|
| Генетическое разнообразие коренных популяций южной Сибири: Филогенетические свидетельства древних контактов С.И. Жаданов, Л.П. Осипова, Л.Э. Табиханова, Т.Г. Шур Некоторые актуальные проблемы современной антропологии |
|
| Изучение полиморфизма гена c-FMS (по DEL425) в популяциях коренных народов Западной Сибири Сенькова Н.А., Осипова Л.П., Ромащенко А.Г. Биологические эффекты малых доз ионизирующей радиации и радиоактивное загрязнение среды: тезисы докладов Международной конференции (Сыктывкар, Республика Коми, Россия, 28 февраля – 3 марта 2006 г.) |
|
| Изучение полиморфизмов DEL425 3'UTR(TC--CA) гена C-FMS в популяциях коренных народов Западной Сибири Сенькова Н.А., Осипова Л.П., Ромащенко А.Г. 13 Международный конгресс по приполярной медицине (Новосибирск, 12-16 июня 2006 г.) |
|
| Профиль функциональной сенсомоторной асимметрии мозга у школьников Ямало-Ненецкого автономного округа Разумникова Е.И., Сенькова Н.А., Осипова Л.П. 13 Международный конгресс по приполярной медицине (Новосибирск, 12-16 июня 2006 г.) |
|
| Оценка репродуктивного потенциала и гормонального контроля половой функции у тундровых ненцев Ямало-Ненецкого АО Осадчук Л.В., Осипова Л.П., Воронич Е.С., Осадчук А.В. Международная конференция «Биорад – 2006». «Биологические эффекты малых доз ионизирующей радиации и радиоактивное загрязнение среды» |
Гранты
| 2011 | Реконструкция процессов антропогенеза: анализ митохондриальных геномов представителей древнего и современного населения Сибири РФФИ, номер гранта 11-06-12006 |
| 2010 | Молекулярные механизмы агрессивного и депрессивноподобного поведения: исследование роли 5-НТ1А, 5-НТ2А и 5-НТ3 серотониновых рецепторов Федеральное агентство по науке и инновациям РФ, номер гранта МК-199.2010.4 |
| 2009 | Применение нейротрофического фактора мозга (BDNF) для коррекции наследственных нарушений функции мозга и поведения Российская Академия Наук, номер гранта 21.1/5.1 |
| Ассоциация между агрессивным поведением и полиморфизмом в генах-кандидатах, регулирующих серотониновую систему мозга РФФИ, номер гранта 09-04-92858-НИСИ-а |
|
| Комплексное исследование генетических, молекулярных и физиологических механизмов депрессии и разработка новых методов ее фармакологической коррекции. Роль наследственных изменений в цитокиновой и серотониновой системах мозга Президиум Сибирского Отделения Российской Академии Наук, номер гранта 18 |
|
| 2008 | Молекулярная биология серотониновой системы мозга в экспериментальных моделях генетической предрасположенности к агрессии и депрессии Российская Академия Наук, номер гранта 6.9 |
| Генофонды и генетическое разнообразие природных и экспериментальных популяций Президиум РАН, номер гранта 26.30 |
|
| 2007 | Роль триптофангидроксилазы-2 - ключевого фермента синтеза серотонина в мозге - в регуляции генетически детерминированного поведения РФФИ, номер гранта 07-04-00209 |
| 2006 | Комплексное исследование генетических механизмов нарушений поведения животных и человека на генном, клеточном, организменном и популяционном уровнях.Роль серотонина мозга Президиум Сибирского Отделения Российской Академии Наук, номер гранта 5.1 |
| Репродуктивные последствия социального доминирования у лабораторных мышей: роль генетических факторов, андрогенного статуса организма и внешних условий формирования иерархии РФФИ, номер гранта 06-04-48937 |
| © 2010-2026 ИЦиГ СО РАН. Все права защищены. |